б) Пластичность инстинктивного поведения насекомых
Различные формы инстинктивного поведения проявляются в отличие от рефлекторных форм только в естественной жизни животных. Тем не менее эти формы поведения не являются более простыми. Инстинктивное поведение часто рассматривают как жесткую, негибкую
47
систему действий, как поведение, не способное к приспособительной изменчивости. Однако это не так.
Перейдем к рассмотрению организации сложных поведенческих актов, в которых сочетаются длинные цепи чисто инстинктивных, жестко организованных автоматизмов и участки условно-рефлекторной индивидуально приобретенной деятельности. Рассмотрим некоторые примеры поведения такого типа. Наиболее явным примером жестко организованного поведения является деятельность насекомых.
Способности пауков ткать свою сеть, пчел строить соты, ос парализовать, но не убивать свои жертвы, предназначая их в качестве "живых консервов" для своего потомства, не являются следствием индивидуального обучения. Появляясь на свет, насекомые, вообще говоря, приносят с собой эту деятельность в готовом виде. Яркой иллюстрацией такого автоматизма является типичное поведение осы при постройке норки для своих детей, рис. 19, а. Схема алгоритма действий в принципе не подлежит коррекции: после снабжения норки пищей и откладки яйца следует процесс замуровывания входа (41; 116-137).
При более детальном рассмотрении эта деятельность выглядит как ряд последовательно включаемых подпрограмм. Сигнал окончания i-го этапа служит пусковым сигналом для начала i+1-го этапа. В частности, несколько начальных этапов состоят в устройстве норки. Их окончание служит сигналом для начала поиска пищи для будущих личинок
 Рис. 19.
Рис. 19. Гибкость инстинктивного поведения; а) часть схемы работ земляной осы по построению норки и заготовлению пищи для потомства. Этапы: 1 - нахождение сверчка, 2 - обездвиживание, 3 - захват добычи и полет к норке, 4 - поиск дальних ориентиров, 5 - поиск ближних ориентиров, 6 - отрывание входа, 7 - проверка состояния норки, 8 - затаскивание сверчка, 9 - оценка количества пищи, 10 - откладывание яиц, 11 - замуровывание входа (по 41). На всех этапах имеют место элементы гибкости и вариативности инстинктивного поведения;
б) различные варианты достраивания личинками ручейника своего домика, частично разрушенного в ходе эксперимента. Пунктир - разрушенные части (по 14)
48
(сверчков строго определенного вида, которых оса не убивает, но парализует, готовя таким образом "живые консервы"). Затем идет этап доставки сверчка к норке, далее этап "проверки" состояния норки - оса входит внутрь для последнего осмотра, оставляя сверчка снаружи. После чего идут окончательные этапы: втаскивание сверчка, откладывание яиц и замуровывание входа.
Во многих экспериментах исследователи после окончания процесса замуровывания на глазах у осы вскрывали вход, вынимали из норки запасы пищи вместе с яйцом и оставляли их около норки. После этого насекомое часто возвращается в норку, проводит там какое-то время, после чего вылезает и снова аккуратно закрывает вход, хотя теперь это не имеет никакого значения. Никогда осы не делают шага назад в алгоритме своих действий и не втягивают пищу и яйцо обратно в норку. Более того, занимаясь заделыванием входа, насекомое часто бегает и наступает на яйцо, не обращая никакого внимания на состояние основной "цели" своей деятельности. Однако результаты работ таких исследователей, как Фабр, Тинберген, Лоренц, и многих других свидетельствуют о том, что при рассмотрении классических примеров инстинктивного поведения насекомых выявляются участки, в которых отсутствует однозначная последовательность действий и проявляется определенная гибкость поведения.
В процессе такого жестко организованного автоматизма имеют место и особые этапы - этапы гибкого поведения. Конечно, эти этапы или участки редки и составляют очень небольшую часть длинных и сложных автоматизмов, но для нашего рассуждения важен сам факт их наличия.
В простейшем случае это способность к повторам последнего действия, которая проявляется по крайней мере в некоторых случаях. Например, в тех же самых экспериментах при подлете к построенной норке оса часто оставляет свою добычу (питание для будущей личинки) у входа, влезает в норку, как бы проверяет ее состояние и затем уже втаскивает добычу. В одном из экспериментов исследователь 40 раз подряд отодвигал добычу от входа в норку, в то время пока оса "проверяла ее состояние". И каждый раз оса вылезала из норки, бежала к добыче, подтаскивала ее к норке и залезала внутрь "с проверкой".
Другой пример гибкости - поиск места входа в норку. В случаях, когда исследователь в промежутке между прилетами осы с добычей к гнезду изменял положение близлежащих ориентиров, оса всегда осуществляла долгое исследовательское поведение, которое, как правило, заканчивалось нахождением входа. (Трудность этой задачи связана с тем, что, улетая в очередной раз за добычей, оса засыпает вход песком, так что при подлете ей приходится полагаться на внешние ориентиры.)
49
Одним из наиболее ярких примеров гибкости отдельных звеньев инстинктивного поведения приводится в работах классика современной науки о поведении животных (этологии) - Нико Тинбергена. Он изучал поведение ос в процессе деятельности по накоплению запасов гусениц для будущего потомства. Оказалось, что если перед проверочным посещением норки исследователь изымал из нее всех накопленных гусениц, то осы затем компенсировали убытки. В итоге после нескольких изыманий некоторые из них приносили в общей сумме по 12-13 гусениц, при нормальном количестве - 5-10 штук. Если же экспериментатор добавлял в норку новых гусениц, то оса реагировала и на это, уменьшая количество приносимой пищи (речь, конечно, идет не о способности к счету, а лишь о примерной оценке части поля зрения занятой добычей).
При этом отмечались и другие аспекты гибкости инстинктивного поведения. В частности, отмечалось, что сигналы, получаемые осой во время проверочного посещения норки, руководят ее деятельностью в течение нескольких дней, несмотря на то, что в этот период она совершает много других типов поведения.
Рассмотрим еще один пример: наличие гибких элементов в ходе инстинктивного автоматизма построения домика-чехольчика личинкой ручейника (рис. 19, б.) Если мы проследим за этапами такой постройки, особенно в условиях, когда экспериментатор частично разрушает домик в процессе его достраивания, то увидим, что даже одна и та же личинка достраивает домик каждый раз по-разному, постройки разных личинок тем более в чем-то отличаются. Естественно, что жесткая и сложная схема инстинкта в основном неизменна: каждый чехольчик имеет определенную длину, состоит из центральной трубки, навеса над входом, днища и крыши, имеет переднюю, заднюю и боковые стороны.
В чем же проявляется гибкость? Животное может научиться строить домик из необычного для него материала, в каком-то смысле изменяя при этом свои строительные движения, способно начинать достраивание после разных степеней разрушения, меняя при этом пропорции частей (14).
Удивительные механизмы гибкости инстинкта, связанные с переключением программ инстинктивной деятельности, демонстрируют пчелы. Если из улья искусственно убрать все группы рабочих пчел, занятых сбором пыльцы и нектара, на эту деятельность переключаются пчелы, занятые совсем другой работой, если убрать всех пчел, строящих соты, то их функции начнут выполнять другие пчелы, занятые до этого сбором меда.
50
Таким образом, гибкость инстинктивного поведения означает всего лишь наличие в жестком автоматизме поведения отдельных мест, допускающих некоторую вариативность действий. Например, действий по достижении результатов некоторого этапа. Набор таких действий, по-видимому, сильно ограничен, различия между ними с точки зрения "инстинктивного существа" отсутствуют.
Моделью гибкости поведения "инстинктивного существа" может быть работа типичной для современной техники схемы - схемы контура управления, например, контура управления процессом наполнения бассейна (рис. 20, а.) Постоянный, заранее заданный уровень воды в бассейне поддерживается путем сравнения в компараторе (от англ, compare - сравнивать) заданного и текущего уровня. При их рассогласовании подается сигнал на включение насоса. В данной простейшей схеме управление осуществляется путем измерения уровня поплавка: опускание поплавка при уменьшении воды в бассейне ниже заданного уровня чисто механически открывает (при помощи известного всем шарового крана) приток воды в бассейн.
Сравним работу этой схемы с работой гипотетического контура управления этапом инстинктивного поведения (рис. 20, б). В качестве примера рассмотрим этап прикрепления строительного элемента (песчинки, палочки, камешка и пр.) при строительстве домика личинкой ручейника. Блок ограниченного набора действий реализует захват строительного элемента любыми лапками, обмазывание его слюной и прикрепление к строящемуся домику. (Ограничениями для работы этого блока могут быть вес и размер строительного элемента, т.е., другими словами, невозможность поднятия элемента или его охвата.)
Блок измерения результатов срабатывает автоматически, например, при наличии мышечного сигнала о возросшей трудности поворотов строительного кирпичика. Работа блока компаратора заключается в сравнении длины домика с длиной тела ручейника и (или) в сравнении мышечных сигналов от пустых лапок и заданных (определенных параметрами тела) сигналов о диапазоне веса кирпичика. В итоге сравнения
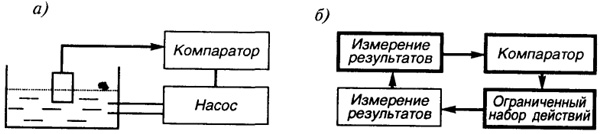 Рис. 20.
Рис. 20. Модель гибкости поведения "инстинктивного существа": а) контур управления процессом наполнения бассейна; б) гипотетический контур управления этапом инстинктивного поведения
51
компаратор посылает сигнал о продолжении или прекращении строительства.
Гибкость данного гипотетического поведения обеспечивается: а) за счет наличия ограниченного набора действий, когда ручейник может использовать любой строительный материал, размеры и вес которого соответствуют размеру и силе его лапок; б) за счет случайности прикрепления строительного материала, что ведет к вариативности формы постройки; в) за счет ограничения точности работы компаратора и т.д.
Принципиальная ограниченность инстинктивного поведения такого рода связана с рядом причин (рис. 21). Во-первых, инстинкт принципиально не способен анализировать внешнюю ситуацию, используя различные зависящие от этой ситуации параметры. Это значит, что в ходе инстинктивного поведения организм не может проводить перебор различных измерительных процедур и не может выбирать из них ту, которая приводит к пригодным в данной ситуации результатам. "Инстинктивное существо" всегда и вне зависимости от ситуации пользуется на определенном этапе вполне определенными, фиксированными действиями, при этом лишь на некоторых участках поведения возможны элементы гибкости.
Эти элементы весьма ограничены и, по-видимому, связаны с такими факторами, как:
- - неразличение (в смысле неточность) действий, когда одинаковые для животного действия могут привести к различным результатам; фактор неразличимости проявляется также в том, что насекомое не способно различить объекты с разными характеристиками и поэтому реагирует одинаковыми действиями в разных ситуациях;
- - возможность повторов некоторых действий (например, действий, определяющих конец этапа), если в результате их проведения отсутствует сигнал о завершении этапа.
Комбинация этих двух факторов может объяснить ранее приведенные факты гибкости поведения насекомых. Например, факты многократного подтаскивания осами добычи к норке или факты определения насекомыми ориентиров при подлете к гнезду.
 Рис.21.
Рис.21. Принципиальные ограничения возможностей инстинктивного поведения
52
Вторая причина ограниченности возможностей инстинктивного поведения, по-видимому, связана с принципиальным отсутствием возможности формулирования целей поведения. Конечно, речь идет о невозможности формулирования произвольных целей поведения, так как можно считать, что все инстинктивное поведение определяется некоторыми не поддающимися изменению целями, сформированными в результате эволюционного процесса.
Третья причина - является прямым следствием второй и связана с принципиальным отсутствием способностей к планированию путей достижения целей. По определению Фабра, "животное ни свободно, ни сознательно в своей деятельности; ... оно строит, делает ткани и коконы, охотится, совершенно не отдавая себе никогда ни малейшего отчета в цели и в средствах. Оно не осознает своих чудных талантов точно так же, как желудок не осознает своей ученой химии" (43; 224-225).
Четвертая причина ограниченности возможностей инстинктивного поведения является следствием всех перечисленных ограничений и заключается в принципиальной неспособности к обучению новым типам поведения. Однако следует отметить определенную сложность проведения границ между новым типом поведения и некоторым вариантом старого, инстинктивного поведения.
В качестве примера можно, например, привести многочисленные данные (14; 132-140) о вариативности строительного поведения личинок ручейника при восстановлении повреждений их домиков, сделанных экспериментатором. После удаления задних двух третей домика личинка № 1 использовала оставшуюся часть домика просто как опору для построения нового, личинка № 3 достроила домик, приделав спереди некоторый кусок, личинка № 6 удлинила трубку спереди и сзади, а также построила новую крышу и т.д.
Возможно, пределы вариативности поведения связаны с критерием, высказанным еще Фабром, который считал, что инстинктивное поведение "умеет справиться со случайностью, лишь бы новое действие не выходило из круга работ, которые занимают его в данный момент" (43; 406).
53
