§ 1. УРОВЕНЬ НЕДИФФЕРЕНЦИРОВАННОЙ ЧУВСТВИТЕЛЬНОСТИ
Самый низший, зародышевый уровень развития психического, отвечающий, согласно А.Н. Леонтьеву, недифференцированной чувствительности, следует непосредственно за допсихической формой жизни и допсихической формой отражения. Соответственно этому фаза фоновой активности (всегда
29
отвечающая, как было показано в § 3 главы 1, предыдущему уровню развития) осуществляется здесь на допсихическом уровне и связана с физиолого-биохимкческими факторами, т.е. может быть названа поисковой лишь условно. К.Э. Фабри отмечает, что на "исходном уровне развития психического отражения первичные формы внешней двигательной активности... детерминировались только раздражимостью".
Можно полагать, что поисковая активность побуждалась на данном уровне прекращением ассимилятивных биохимических реакций вследствие происходившего по тем или иным причинам отделения живого организма от содержащих жизненно необходимые свойства или компоненты частей среды. В простейшем случае (при незначительном пространственном разъединении) она, по-видимому, выражалась в беспорядочных хаотичных движениях, в конце концов приводивших к требуемому для собственно активной фазы деятельности сближению с предметом потребности. Эти движения, по всей вероятности, имели либо биохимическую, либо физико-химическую природу.
Очевидно, что первичная недифференцированная чувствительность направлена на необходимые для поддержания жизни свойства среды, приобретающие сигнальный характер. Собственно активная фаза деятельности проявляется в реакции на эти свойства, наступающей при определенном (на первых этапах развития, по-видимому, очень тесном) сближении с предметом потребности. По его достижении все как бы возвращается на круги своя, реакция (R) фактически знаменует восстановление внутренней деятельности. Внешняя деятельность является здесь как бы простым придатком внутренней (ассимилятивных процессов), составляющей на уровне недифференцированной чувствительности основную, существенно преобладающую часть жизнедеятельности.
Итак, общую структуру жизнедеятельности на уровне недифференцированной чувствительности можно представить в следующем виде: актуализация потребности (прекращение ассимилятивных процессов) — фоновая активность — стимул-реакция (S-R) — достижение предмета потребности (восстановление ассимилятивных процессов).
Стимул — реакция (S-R) выступают как прообраз рефлекторного акта ("прарефлекс"), который состоит из непосредственно связанных афферентного и эффекторного звеньев. Отметим, что собственно активная фаза внешней деятельности осуществляется лишь на границе с внутренней, в "прарефлексе".
30
Психическое отражение на данном уровне выступает как чувствительность к жизненно значимым свойствам среды. Она является и внутренней стороной результата отражения. В то же время уже здесь можно выделить и внешнюю сторону результата отражения — свойства среды, имеющие сигнальное значение.
Среди современных живых организмов не встречаются животные, отвечающие уровню недифференцированной чувствительности в его первоначальном виде. К.Э. Фабри указывал, что "строение и поведение современных "простейших" являются уже намного более сложными, чем у их ископаемых предков. Он полагает, что при зарождении животной жизни существовал более низкий, чем у современных одноклеточных, действительно первоначальный уровень развития.
В то же время многие современные животные близки к уровню недифференцированной чувствительности. Это простейшие (амебы, инфузории, жгутиковые и т.д.), многие низшие многоклеточные (губки, коралловые полипы и др.), для которых К.Э. Фабри выделил низший уровень стадии элементарной сенсорной психики.
Выше отмечалось, что на первоначальном уровне психической (животной) жизни фаза фоновой активности еще не связана с психикой и остается обусловленной допсихическими формами отражения. Поведение современных простейших подтверждает указанный вывод.
Способы передвижения простейших весьма разнообразны. Некоторые пассивно перемещаются вместе с толщей воды, в которой они находятся, есть простейшие, перемещающиеся реактивным способом. Амебы двигаются посредством перераспределения плотности протоплазмы, как бы "переливая" ее из одного участка тела в другой. Большинство же простейших передвигается с помощью жгутиков или ресничек, представляющих собой волосовидные плазматические выросты. Однако, как почеркивал К.Э. Фабри, независимо от перечисленных способов, движения простейших (кинезы) в основном определяются "непосредственным воздействием градиентов интенсивности биологически значимых внешних факторов", таких, как химический, температурный и т.п. С какими-либо психическими процессами локомоция (передвижение) простейших в фазе фоновой активности не связана.
Для простейших характерны два вида кинезов — ортокинез и клинокинез. При ортокинезе животные осуществляют поступательное движение с переменной скоростью,
31
определяемой градиентом воздействующего внешнего фактора. При клинокинезе изменяется направление движения. Изменение направления движения совершается хаотично, так что животное "находит" наиболее благоприятный вектор движения после "проб и ошибок".
Кинезы носят ненаправленный характер и не обеспечивают подлинной ориентации. Ориентирующие процессы, вносящие адекватную коррекцию в хаотическое изменение характера движения, называются таксисами.
Таксисы простейших, как и их кинезы в целом, не связаны с проявлением психического. Показательна в этом отношении тенденция к движению вверх инфузории туфельки, не имеющей гравитационной чувствительности. Ориентация движения туфельки определяется действием содержимого пищеварительных вакуолей, которое давит на их нижнюю часть. Если над туфелькой, проглотившей порошок железа, поместить магнит, она начинает двигаться не вверх, а вниз: порошок давит не на нижние, а на верхние части вакуолей, что приводит к обратной ориентации движения.
Не связаны с проявлением психики и реакции простей ших на температуру. К.Э. Фабри отмечает, что "здесь не посредственно необходимая для поддержания жизни энер гия еще тождественна опосредствующей энергии, сиша лизирующей о наличии жизненно необходимого компонента среды". Какие-либо специфические терморецепторы у простейших отсутствуют. К.Э. Фабри полагает, что реакции простейших на температуру сходны с биохимическими реакциями типа ферментативных процессов.
Собственно активная фаза жизнедеятельности простейших, связанная с зачаточными проявлениями психического, осуществляется в непосредственной близости от предмета потребности. По имеющимся данным, амеба реагирует на пищу с расстояния 20—30 мкм. Активно реагировать на предмет пищевой потребности способны и хищные инфузории. К собственно активной фазе деятельности простейших относятся и фобические реакции, проявляющиеся, например, у амеб и инфузорий. Так, при контакте с твердым предметом либо оказавшись в каких-то других неблагоприятных условиях туфелька прекращает движение и "сьеживается", что обычно предшествует выделению стрекательных капсул.
Пластичность поведения — способность к индивидуальному изменению врожденного видотипичного поведения —
32
у ростейших выражена весьма слабо. Она ограничивается наиболее примитивной формой — привыканием. В опытах йшецкого ученого Ф. Брамштедта парамеции определенное время держались в сосуде треугольного или квадратного сечения, а затем перемещались в более крупный сосуд круглого сечения. В нем они продолжали сохранять привычное движение по траектории, повторяющей периметр прежнего сосуда (рис. 1.3).
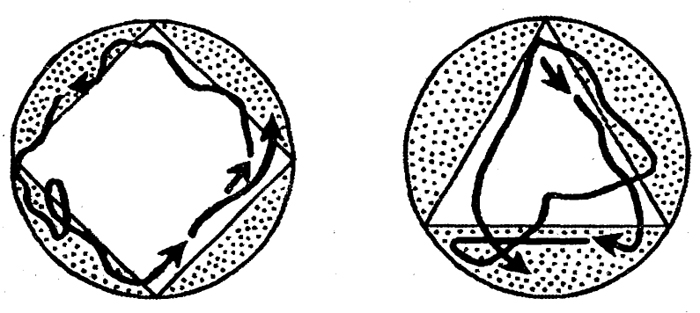
Рис. I.3. Привыкание у инфузории туфельки (по Ф. Брамштедту)
33
