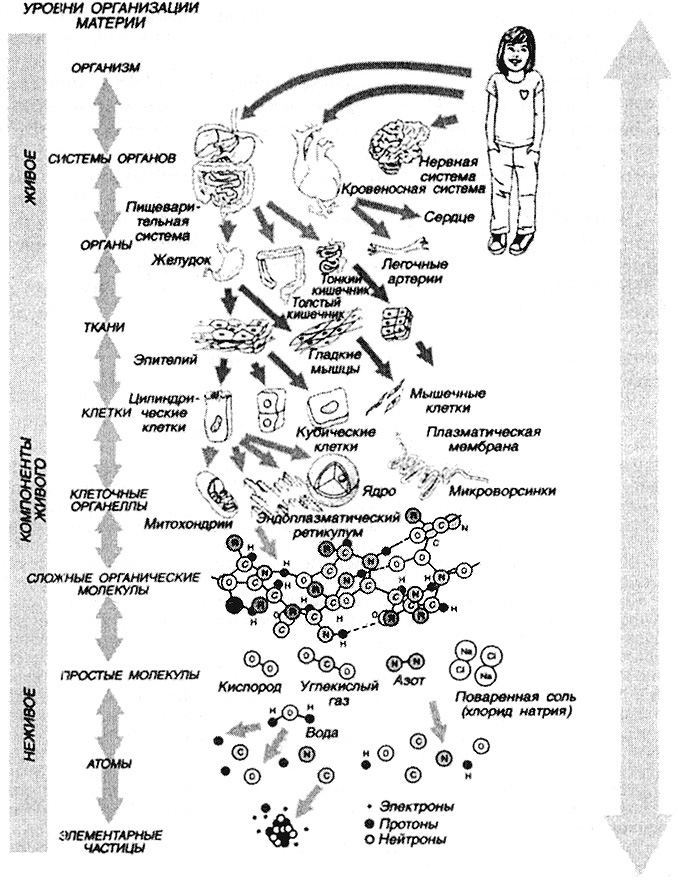
Жизнь, как ее определяют многие энциклопедические издания, являет собой одну из форм существования материи, закономерно возникающей при определенных условиях в процессе развития материи. Это определение отнюдь не является исчерпывающим. Как известно, жизнь - одна из основных тем как философского размышления на протяжении всей человеческой истории, так и предмет естествознания и теологии.
По образному выражению французского естествоиспытателя Ж. Кювье, "Жизнь представляет... более или менее сложный вихрь... направление которого постоянно и который всегда захватывает молекулы, обладающие определенными свойствами... в него постоянно проникают и из него постоянно выбывают индивидуальные молекулы... Пока это движение существует, тело, в котором оно имеет место, живо, оно живет. Когда движение окончательно останавливается, тело умирает" (А.А. Горелов, 1998, с. 48).
Процесс божественного сотворения рассматривается как совершившийся одномоментно и всего однажды. В этом случае естественнонаучное изучение возникновения жизни становится невозможным, хотя в теологии оно продолжается. Теория самопроизвольного и спонтанного возникновения жизни была известна в древних цивилизациях Китая, Египта и активно рассматривалась Аристотелем. В основу этой теории положена идея "активного начала", которое присутствует во всех живых организмах и распространяется на неживые предметы. В XVII-XVIII вв. благодаря трудам крупнейших ученых А. Левенгука, Л. Пастера, Ф. Леди и других пришли к отвергающему теорию активного начала выводу, что жизнь может возникнуть только из предшествующей жизни. Теория самопроизвольного возникновения жизни была отвергнута концепцией биогенеза. Имеется еще достаточно большое количество и других гипотез возникновения жизни, но и как упомянутые выше они не отвечали на вопрос: откуда взялся самый первый живой организм.
Разработанная рядом ученых теория стационарного состояния отвечает на этот вопрос однозначно: жизнь была всегда и не возникала вновь, а проявлялась в форме вечно бессознательного существования. Но здесь есть одно важное допущение, которое эту теорию опровергает. Если жизнь существовала всегда на Земле, то и Земля существовала всегда, но этот факт, как известно, неверен. Теория панспермии, утверждающая, что жизнь занесена на нашу планету извне, в целом возникновения жизни не объясняет, а просто выносит проблему за пределы Земли.
Признаваемая подавляющим большинством современных биологов теория биохимической эволюции полагает, что возникновение жизни и развитие многообразных жизненных процессов связано с различными зачастую сложными биохимическими и физическими реакциями. Жизнь возникла в результате превращений и переходов от сложных органических веществ к простым живым организмам. Согласно данным современной науки, в процессе соединения возникших абиотическим путем аминокислот образовалась некая материальная система, составленная из управляющей и управляемой частей (ядро клетки и цитоплазмы). Ядро клетки содержит молекулы дезоксирибонуклеиновой кислоты - ДНК, в которой заключен информационный "генетический" код. Данный код в виде порядка расположения четырех оснований, соединяющих две цепочки атомов, определяет последовательность всех процессов жизнедеятельности организмов - биосистем, где осуществляется обмен веществ с окружающей средой, в ходе которого организм получает все необходимые вещества для обеспечения его роста и развития, а также размножения в виде образования дочерних организмов.
Наиболее проработанной и поддерживаемой современными учеными среди огромного множества имеющихся теорий и гипотез, объясняющих возникновение и развитие его органического мира, является теория эволюции. Вклад в нее сделали Ламарк, Дарвин, Уоллес, а дополнения, внесенные достижениями современной генетики, палеонтологии, молекулярной биологии, позволяют сейчас определить теорию органической эволюции жизни как путь естественного отбора детерминированных генетических признаков (рис. 30).
Начиная с XIX века изучение жизни связывается не только с классической естественной и теологической проблематикой, но и с так называемой неоклассической, разработанной в рамках современной философии. Но это уже предмет иной науки, нежели той, что рассматривается в данной книге. Однако, на наш взгляд, следует высказать некое важное утверждение: экологическая наука имеет предметом своего изучения живые организмы, одним из которых является человек, существо разумное, способное собственно и изучать... Это и есть один из важных постулатов современной экологии как мировоззренческой науки: "Экология изучает объект в виде живого организма как проявления
жизни, связанной со средой не только условиями обитания, но и условием своего возникновения. В этом случае нарушения во взаимосвязях чреваты разрушением объекта изучения - живого организма, а значит и собственной жизни". Выходом является учет в выборе
тенденций развития, наличие у современной жизни человека, как единственного носителя Разума и Души человечества, как носителя Коллективного Разума и Души...
В.И. Вернадский писал: "На земной поверхности нет химической силы, более постоянно действующей, а потому и более могущественной по своим конечным последствиям, чем живые организмы, взятые в целом... Все минералы верхних частей земной коры - свободные алюмокремниевые кислоты (глины), карбонаты (известняки и доломиты), гидраты окиси Fe и А1 (бурые железняки и бокситы) и многие сотни других - непрерывно создаются в ней только под влиянием жизни". Во многом поверхностная часть, довольно мощная часть земной коры и ее внешний облик сформирован жизнью.
Прекрасно известно, что геологическое время весьма отлично от обыденного ощущения и понимания времени человеком, но это понимание показывает, что продолжительность действия жизни на "слои земные" соизмерима со сроком существования земной коры, с ее абсолютным возрастом.
Рассмотрим в виде краткого описания глобальную эволюцию Земли (по В.И. Дубинину, 1997). Начиная с 40-х годов нынешнего столетия, с работ акад. О.Ю. Шмидта, сложилась вполне обоснованная по уровню современного научного знания гипотеза образования Земли. Основное представление о формировании планет, составляющих Солнечную систему, заключается в том, что они образовались за счет слипания и дальнейшего роста (аккреации) твердых частиц из протопланетного пылегазового облака, окружавшего Солнце.
По расчетам ряда современных специалистов рост Земли не был постоянно ускоряющимся, а носил циклический характер, вначале происходило довольно быстрое формирование массы Земли, которое по мере исчерпания твердого вещества в протопланетном облаке замедлилось и трансформировалось в уплотнение собственно планеты Земля. По данным B.C. Сафронова, при аккреации Земли выделилось гигантское количество гравитационной энергии - около 23,3 × 1038 эрг. Эта энергия была так велика, что прото - Земле грозило не только переправление первичного вещества, но и полное его испарение. Однако большая часть энергии выделялась в приповерхностных зонах и терялась в окружающем пространстве в виде теплового излучения. За 100 млн. лет сформировалось около 99 % массы современной Земли. Процесс аккреции Земли сопровождался попеременным разогреванием и остыванием вещества, что обусловило на первичном этапе ее существование в виде недифференцированной планеты без ядра и земной коры.
По современным представлениям Земля после своего формирования была относительно холодным телом однородного состава с температурой ниже необходимой для переплавления.
Вся дальнейшая эволюция Земли как планеты была связана с качественным составом ее вещества, полученной энергии при образовании и взаимодействиями с ближайшим космическим телом - Прото-Луной. Именно эти факторы обусловили эндогенные процессы, проявившиеся за счет энергии распада радиоактивных элементов и гравитационной дифференциации исходного вещества Земли. По своим параметрам и свойствам поверхностные слои тела Земли соответствовали реголиту - мелкопористому веществу, образованному из постоянно оседающей тонкодисперсной пыли, непрерывно обновляющейся
104
при взрывных воздействиях падающих на Землю планетезималей - агрегированного космического материала. При этом процессе происходило выделение воды и углекислого газа, которые мгновенно связывались ультраосновным по составу земным веществом - реголитом. Общий теплозапас Земли определялся главным образом скоростью ее роста, но величина тепла Земли была недостаточной для полного переплавления ее вещества, что существенно отличало ее от ближайшего спутника - Прото - Луны и привело за 4 млрд. лет к формированию земного ядра. Этот процесс продолжается и сейчас за счет гравитационной дифференциации мантийной составляющей планеты.
На начальных этапах примерно 600 млн. лет в катархее в криптотектоническую эпоху происходил медленный разогрев земных ее недр, который дополнялся приливными воздействиями Луны. Тектономагматическая активность смогла проявиться только тогда, когда температура в недрах Земли поднялись до возможности переплавления земного вещества. Примерно 4 млрд. лет тому назад Земля сформировала астеносферу, что в сочетании с приливными взаимодействиями привело к ускоренному отдалению Луны от Земли. Очень важные процессы происходили в это время на Луне, которая собирала из околоземного пространства все малые космические тела и испытывала активную бомбардировку своей поверхности, что привело в конце концов к разрушению ее анортизитовой коры и активным излиянием базальтовых расплавов.
По данным В.Е. Хаина, эти два события - начало тектонической деятельности на Земле и проявление базальтового магматизма на Луне взаимно связаны и привели, в частности, в конце концов к выделению и росту в центре нашей планеты плотного оксидно-железного земного ядра. В течение первых 600 млн. лет Земля обладала разреженной атмосферой, состоящей из благородных газов и отличалась полным отсутствием гидросферы.
По В.А. Зубакову (1997), ландшафты на поверхности Земли в самом начале ее существования были по нашим представлениям весьма мрачными. Это была суровая неприветливая холодная пустыня с черным небом с немерцающими яркими звездами и одновременно слабогреющим желтым Солнцем. Появляющаяся Луна была огромным ярким диском без пятен морей. Вся поверхность Земли была покрыта кратерами, которые под действием непрерывных приливных землетрясений были постепенно сглажены и перекрыты мощным слоем реголита. В целом поверхность Земли мало изменялась, только в экваториальной зоне поверхностное вещество было раскаленным из-за ливневого выпадения каменных и железных обломков из спутниковых роев в околоземном пространстве.
Солнце пересекало небосвод за 3 ч, чтобы через 3 ч снова подняться над горизонтом. Движение Луны было медленнее, так как она вращалась в ту же сторону, что и Земля, а фазы Луны сменялись всего за 8 ч. Видимая площадь диска Луны была в 300-350 раз больше современной, и так как Луна была горячей планетой, то диск светился темно-красным цветом и даже обогревал Землю. Луна вращалась вокруг Земли по орбите с радиусом примерно 25 тыс. км (сейчас радиус лунной орбиты составляет примерно 384,4 тыс. км). Это в сочетании с большими размерами Луны приводило к тому, что амплитуда лунных приливов достигала 1,5 км. Лунные же приливы следовали за Луной и каждые 18-20 ч потрясали земной ландшафт. К концу катархея лунные приливы снизились до 7 м и сейчас составляют в твердой Земле вблизи подлунной точки 45 см. Быстрое отдаление Луны от Земли в катархее обусловило невозможность перегрева и пере плавления Земли и привело к постепенному снижению приливной сейсмичности. Следует заметить, что сейсмичность носила в это время исключительно экзогенный характер, так как никакой тектонической деятельности на Земле еще попросту не было. Земное вещество было практически стерильным за счет воздействия жесткого космического излучения, а также было лишено летучих форм, которые, если и образовывались, то сразу же сорбировались поверхностью высокопористого реголита и медленно перемещались в ее недра. Газы же устранялись активным излучением молодого Солнца.
В наступившем архее на Земле начали проявляться более сложные внутренние эндогенные процессы, которые явились следствием гравитационного разделения земного вещества с выделением ядра, силикатной мантии и астеносферы, активизацией радиоактивного
105
распада элементов в приповерхностных зонах и продолжавшимися приливными взаимодействиями с Луной.
В.А. Зубаков, ссылаясь на работы ряда специалистов и идеи В. Эльзассера, утверждают, что начало дифференциации земного вещества было положено путем выплавления из него металлического железа на границе верхней мантии. В целом за счет реализации приливной энергии сформировалась довольно своеобразная структура Земли. Распространявшаяся от экваториальной зоны зона тяжелых расплавов железа и его оксидов перекрывала холодное вещество умеренной плотности (плотность отличалась почти в 1,5-2 раза). Такая структура была гравитационно неустойчивой, что должно было привести к разрушению системы, чего, однако, не произошло. Холодная сердцевина постепенно выдавилась к одному из полюсов Земли, а стекание тяжелых железных расплавов осуществлялось через противоположный полюс, чему способствовало то, что главная ось момента инерции Земли совпадала с осью ее вращения.
Примерно 2,7-2,6 млрд. лет тому назад в конце архея за весьма короткий период времени у Земли сформировалось плотное ядро и это же время характеризуется исключительной тектонической активностью. Примерно тогда же все первозданные ландшафты Земли были разрушены, вся первичная литосфера погрузилась в недра горячей мантии, началось деформирование первичных тектонических структур. Перепад высот на земной поверхности был весьма´небольшим-до 1,5-2,0 км. К концу архея сформировался мелководный мировой океан за счет образовавшейся в магмагенных процессах воды. Луна уже в меньшей степени оказывала приливное воздействие, так как удалилась более чем на половину от современного расстояния, хотя лунные приливы высотой около 0,5м деформировали земную поверхность примерно каждые 5 ч. Скорость вращения Земли была еще достаточно большой, сутки были равны примерно 19 ч, а в году было 460 суток.
Неблагоприятные условия для развития жизни на Земле сохранились практически до начала протерозоя, т. е. до тех пор, пока не начал действовать процесс дегазации земного вещества. В самом начале этого процесса, еще в конце катархея, большая часть формировавшейся воды и других элементоорганических соединений поглощались реголитом, но при этом его высокая пористость и исключительная сорбцирнность создавали определенный благоприятный фон для образования сложных органических соединений. Некоторую роль здесь играло наличие в реголите и вулканических пеплах в значительных количествах и свободном состоянии таких элементов, как хром, железо, кобальт, никель, свинец, платина, которые, как известно, обладают каталитическими свойствами в процессах синтеза органических веществ. Таким образом, по представлениям В.А. Зубакова, на базе идей В.И. Вернадского и И. Пригожина в земной истории примерно 4 млрд. лет тому назад в результате контакта с Прото - Луной при поглощении Землей расплавленного ядра своего спутника нашей планетой кроме значительного объема вещества была получена дополнительная энергия для дифференциации геооболочек, для появления сильного геомагнитного поля и атмосферы в виде плотной и мошной газовой и паровой оболочки. Благодаря ей на Земле появилась свободная вода, насыщенная биогенами - Н, С, N, О, Р, S, которые выделились за счет перемыва реголита (вещества поверхности планеты), сформированного, в частности, и за счет выпадения космической пыли.
По мере дегазации Земли и развития гидросферы, возникшие в реголите примитивные формы жизни в виде простых ассоциаций сложных органических молекул или образований, содержащих рибонуклеиновые кислоты, смогли быть перемещены в воду сформировавшегося Мирового океана. Эта вода, образно говоря, была "животворной". Именно в ней осуществилась, по предположению В.А. Зубакова, "хиральная" революция - революция разделения вещества на элементарные составляющие жизни - накопление левоориентированных стереоизомеров аминокислот и правоориентированных сахаров. Дальнейшее совершенствование жизни должно было происходить по биологическим законам развития живой материи, таким, как "фильтрующие" свойства внешней среды и направленное гравитационное давление.
В обычных космических условиях, по данным исследований метеоритов, "левые" и
106
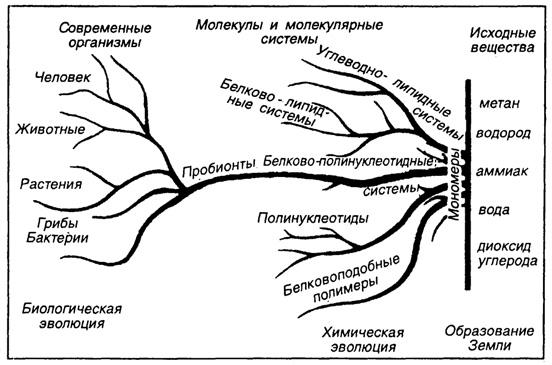
Рис. 31. Переход химической эволюции в биотическую
"правые" стереоизомеры существуют в примерно равном соотношении. На Земле же возникла определенная дисимметрия в их содержании, что собственно и послужило толчком для дальнейшей эволюции элементов жизни. По гипотезе А. Кернс-Смит (В.А. Зубаков, 1997) такая дисимметрия явилась следствием накопления аминокислот в дефектах кристаллической решетки глинистых минералов. Глинистые же минералы обладают магнитными свойствами и в процессе своего образования при выветривании первичных полевых шпатов были ориентированы по магнитному меридиану. Так что дисимметрия образовалась за счет магнитного взаимодействия между Землей и Прото - Луной. Данный факт ранней земной истории был началом изменений в химической эволюции земного вещества, особенно в ее поверхностной зоне. Химическая эволюция разделилась на две "руки" ("хирос" - по-гречески - рука) - гидрогенные осадочные минералы, которые отличаются вполне четкой внутренней симметрией - полимеры на углеродной основе, дисимметричные по своему строению. Далее последовало разделение на нуклеотиды и белки, появление генов, образование клетки и так далее до формирования собственно биосферы. Естественно, высказанное беглое рассуждение не описывает ни длительности, ни мучительности развития жизни на Земле (рис. 31).
В катархее и архее, на отрезке времени между 4,0-2,8 млрд. лет тому назад, когда сформированная к этому времени атмосфера значительно уплотнилась и насытилась различными парами за счет исключительно активной вулканической деятельности. Это привело к тому, что солнечный свет не достигал поверхности Земли. Высокая плотность атмосферы даже при весьма низкой светимости Солнца* (примерно в 4 раза меньше современного) привела к формированию высокого парникового эффекта. Следствием этих факторов стал ровный, очень теплый климат с температурами в диапазоне 200-400° С. Мелкие, пересыщенные солями обширные моря были сосредоточением проявлений жизни. Органический мир был представлен, как пишет В.А. Зубаков, доядерными бессмертными микроорганизмами - прокариотами. Прокариоты были гетеротрофными архебактериями, дышащие метаном и производными серы. Прокариоты не обладали оформленным клеточным ядром и типичным хромосомным аппаратом. Наследственная информация реализовывалась через ДНК.
При ослаблении вулканизма в конце катархея и особенно на последних стадиях архея, примерно на этапе 3,0-2,5 млрд. лет тому назад, произошли существенные
107
изменения в атмосфере Земли, которая стала значительно тоньше и несколько прозрачнее. В еще весьма облачной атмосфере появилось Солнце и хотя его светимость несколько увеличилась, но отсутствие "обогрева" за счет вулканизма привело к практическому исчезновению парникового эффекта. Следствием стало существенное похолодание климата, температура снизилась сначала до 80-50° С, а затем и вовсе до 10-4° С. Моря и океаны увеличились по площади и стали значительно глубже, на больших глубинах установилась весьма постоянная, но достаточно низкая температура. На материках, особенно в высоких широтах Земли, началось первое в геологической истории покровное оледенение, продолжавшееся около сотни миллионов лет.
В этих, уже достаточно суровых, относительно прежних, условиях произошли определенные изменения в органическом мире Земли. В дополнение к архебактериям и эубактериям появились автотрофные цианобактерии. Эубактерии или эукариоты, которые к этому времени получили достаточное развитие, представляли собой уже оформленные клеточным ядром образования с отделенной от цитоплазмы ядерной оболочкой. Генетический материал был сформирован в хромосомах. В клетках уже имелись митохондрии, пластиды и другие органоиды. Появление же цианобактерий, использовавших для дыхания азот, привело в конечном счете к постепенному вытеснению архебактерий и эубактерий, что в результате эволюции сделало цианобактерий жизненными монополистами. По В.А. Зубакову (1997), цианобактерий имели очень важную особенность, чрезвычайно существенную для дальнейшего развития органического мира. Этой особенностью было то, что при поглощении азота они выделяли в окружающую водную среду кислород, который в целом был продуктом их жизнедеятельности. Для существовавших же на Земле архебактерий кислород был смертельным ядом. Здесь следует сделать некоторое геологическое отступление.
Геологические данные говорят о том, что фотосинтезирующие одноклеточные микроорганизмы, водоросли, способные окислить железо, появились еще в раннем архее (около 3,8 млрд. лет тому назад), подтверждением чего является наличие железорудных формаций, сложенных окислами трехвалентного железа, в частности, это формация Исуа в Западной Гренландии.
Начиная с архея, масса свободной воды на Земле была сравнительно мала, что позволяет достаточно условно называть имевшиеся тогда моря "мировым океаном". Породы земной коры подвергались гидратации и углекислый газ весьма слабо связывался в карбонатах, что обусловило повышенное парциальное атмосферное давление.
В атмосфере было весьма мало кислорода, хотя отмеченные выше цианобактерий уже производили его в заметных количествах. Дело в том, что произведенный ими кислород, а также кислород, попутно появившийся в других химических реакциях, быстро связывался свободным (металлическим) железом в мантийном веществе, которое активно выносилось на поверхность планеты в рифтовых зонах и реагировало с компонентами атмосферы и гидросферы. Масса произведенного кислорода по расчетным оценкам соответствовала или была даже меньше способного к связыванию его железа, выносимого из недр. Атмосфера в раннем протерозое (примерно 2 млрд. лет тому назад) состояла только из азота, малого количества водяного пара, аргона и сотых долей процента углекислого газа (оксид углерода полностью отсутствовал).
Резкое падение парциального давления углекислого газа вызвало глобальные климатические изменения: исчез парниковый эффект и температура на поверхности Земли резко упала, наступил указанный выше первый на Земле ледниковый период.
Главным же фактором в эволюции жизненных процессов стал именно процесс перехода атмосферы к нейтральному состоянию относительно активности углекислого газа. Это как бы подхлестнуло в развитии эукариоты, которые стали приобретать новые формы одноклеточных организмов и водорослей с выделенным обособленным ядром и другими внутренними структурами.
Металлическое железо исчезло из верхних зон мантии лишь на границе протерозоя и фанерозоя и это сразу же привело к тому, что механизм связывания кислорода с двухвалентным гидроксидом железа перестал действовать. Кислород стал накапливаться
108
в атмосфере и его парциальное давление стало резко повышаться. Этот процесс продолжался почти до конца протерозоя - венда.
Необходимо остановиться также на том факте, что фитопланктон объемом своей биомассы во многом обязан растворенным в воде соединениям фосфора. Геологическими данными установлено, что концентрация фосфорных соединений была всегда в равновесном состоянии относительно базальтов океанической коры и практически равной современной. Это значит, что в протерозое биомасса фитопланктона в океане соответствует современной. Весьма важным экологическим обстоятельством следует также признать, что в связи с отсутствием в протерозое, да и в самом начале палеозоя, наземных растительных форм захоронение органического углерода в форме углей или углеводородов обеспечивает накопление кислорода в атмосфере. Этот процесс в протерозое был весьма мощным, так как в океанах господствовала восстановительная среда, поэтому осадконакопление из фитопланктонного материала проходило без окисления.
Однако пока не прекратился процесс "подачи" железа из рифтовывх зон,практически весь кислород был обречен на связывание. Когда же около 2 млрд. лет тому назад запасы активного железа были практически исчерпаны, то на Земле свершилась геохимическая революция - с этого момента земная атмосфера и океаническая гидросфера стала превращаться из восстановительной в окислительную. Кислород, по тем понятиям, стал отравлять окружающую среду - случился первый - прерифейский - глобальный экологический кризис, пик которого пришелся примерно на время 1,7-1,8 млрд. лет тому назад.
Как пишет В.А. Зубаков, этот кризис отчетливо фиксируется появлением в материковых отложениях ожелезненных красных песчанников, что свидетельствует о том, что в атмосфере количество кислорода достигло "точки Юри" - одной тысячной доли современной концентрации.
Самым первым следствием кризиса была гибель подавляющего числа архебактерий в земном океане. Из их остатков сформировались огромные скопления нефти, газа, графитов в отложениях рифея. Вторым важным следствием стало разделение биосферы на две принципиально отличные по жизненным принципам ветви, кстати весьма неравнозначные по объему. Исходная биосфера восстановительного характера уцелела только в донных, преимущественно глубоких зонах, где сформировались локальные резерваты хемобиосферы. На земной поверхности воцарилась оксибиосфера, при этом освободились значительные по объемам экологические ниши. Дальнейший процесс развития новых форм жизни изучен еще слабо, наибольшую обоснованность в его описании имеет теория эндосимбиоза развития по Л. Маргулис. В.А. Зубаков (1997) так описывает ее основные положения: в результате многоэтапных коопераций доядерных органелл с гипотетическим протоядерным организмом уркариотом (по-гречески - творящийся ядерный) возникло новое направление в жизненных формах. Вначале появилась вышеназванная крупная ядерная клетка - эукариот, способная дышать кислородом и генетически активная, совершенствующаяся путем мутаций. Именно генетическая активность резко усилила эволюционный процесс, а главное появилась возможность бесконечного усложнения жизненных форм. И, в частности, кроме всего прочего, жизнь на Земле невозможна без бактерий особых форм, которые определяют завершенность жизненных циклов, а именно выполняют важнейшую функцию деструкции органического материала, синтезируемого эукариотами.
В раннем палеозое (кембрии и ордовике) в водной среде океанов происходило постепенное усложнение жизненных форм, дышащих кислородом, растворенным в воде. От одноклеточных, затем многоклеточных мягкотелых - медуз, червей и им подобным и, наконец, скелетных - раковинных моллюсков и рыб. Дальнейшее повышение парциального давления кислорода в земной атмосфере и растворенного в воде кислорода в гидросфере привело к тому, что в начале фанерозоя были заложены генетические основы для возникновения и дальнейшего совершенствования практически всех типов скелетных и бесскелетных животных, а также к завоеванию и развитию жизненных форм на суше и совершенствованию форм растительности и эволюции микроорганизмов. В геологическом отношении на протяжении фанерозоя земная поверхность переживала
109
активные процессы трансгрессии и регрессии океана. В это время формировались и распадались древние материки от остатков Пангеи до образования и последующих преобразований Северной Лавразии и Южной Гондваны, и до практически современного положения и конфигурации материков. Глобальные движения в земной коре имели значительное влияние на эволюцию жизненных форм, но это не является предметом нашего изучения и поэтому мы остановимся только на общих характеристиках эволюции биосферы на нашей планете вслед за описанием, данным В.А. Зубаковым (1997).
К середине палеозоя - в силуре, девоне содержание кислорода в атмосфере достигло почти 10 % от современной концентрации. Одновременно произошли и другие изменения в атмосфере, в частности, проявилась стратификация, сформировался озоновый экран как результат сложнейших химических реакций с образованием аллотропной модификации кислорода О3 (озона). В это время происходило активное освоение жизненными формами земной суши. Первыми завоевателями ее были водоросли и древние, еще бесскелетные рыбы, которые из океана и морей сначала перебрались в озера и реки, где сформировали скелет и плавники, а затем в виде кистеперых рыб, перебираясь из водоема в водоем, заложили возможность образования амфибий, оснащенных легкими и лапами. Из растений на суше появились первые высшие формы - примитивные риннофиты. Это в общем-то была новая знаковая точка в эволюции - были созданы предпосылки к цефализации животного мира.
В позднем палеозое отмечается значительный расцвет жизненных форм на суше как среди растений, так и сухопутных позвоночных. На границе перми и триаса, т. е. на переходе от палеозоя к мезозою, почти целиком исчезли четырехлучевые кораллы, трилобиты, фузулиниды и появились гастроподы, двустворки, рептилии. Последние заняли господствующее положение среди животных на суше. В морской фауне преобладали белемноидеи, аммоноидеи и др.
Очень значительный, носящий катастрофический характер, рубеж в эволюции биосферы отмечается на границе мезозоя и кайнозоя (между меловым и палеогеновым периодом). Большое количество групп животных вымирает почти полностью (аммоноидеи, белемноидеи, а также динозавры, ихтиозавры и другие рептилии), появляются новые моллюски и явный толчок в развитии получили млекопитающие. Значительные изменения произошли и во флоре, особенно среди покрытосеменных. Половина всех родов к началу палеогена вымерла, причем это случилось настолько быстро, что по мнению многих исследователей не может быть объяснено исключительно факторами развития Земли и поэтому были сделаны попытки найти причины катастрофы в окружающем Космосе. Естественно, отрицать факт влияния космических факторов на развитие Земли полностью нельзя, но нам представляется, что по мере накопления геологических и палеонтологических знаний многое будет объяснено земными причинами.
Действительно, изменение пространственного расположения, размеров и формы как континентов, так и океанов оказывало существенное воздействие на мощь и структуру океанских течений, на распределение и характер сезонных колебаний температуры, а следовательно, и на распределение климатической зональности и биологической продуктивности, т. е. на формирование экосистем.
Подавляющее большинство (до 90 %) всех видов морских животных обитает на континентальных шельфах или на мелководьях до 200 м. Разнообразие и количество морской фауны уменьшается по мере увеличения широты, до полярных регионов, где оно почти в 100 раз меньше, чем на тропическом шельфе. Степень биоразнообразия четко коррелирует с изменением устойчивости пищевых запасов, которые связаны с климатом. Используя метод актуализма, становится возможным на основании глобальной модели конечной кинематики главных литосферных плит, а также данных о генеральных изменениях уровня Мирового океана и климатических трансформаций объяснение характера изменений числа семейств как в мелководной, так и в определенной мере, глубоководной и наземной фауне и отчасти флоре. На этих палеореконструкциях географии и климата возможно вполне четкое объяснение массовой гибели многих видов на границах палеозоя и мезозоя, мезозоя и кайнозоя.
110
Наряду с климатическими факторами значительную роль играли глобальные и региональные химические изменения степени солености и качественного состава вод Мирового океана. Например, хорошо известна массовая и быстрая гибель многих кораллов на океанических островах в начале позднего мела. Но именно в это время отмечена последняя из известных глобальная трансгрессия, которая привела к тому, что карбонаты из океана наполнили мелководные значительные по площади континентальные моря и к перекрытию ими отложений мелового периода. Уменьшение концентрации карбоната кальция в океане разрушило океанические экосистемы для многих моллюсков, например, рудистов.
Палеомагнитные реконструкции, т. е. установление истории развития и трансформаций магнитного поля, показывают также на весьма ощутимую его роль в биологической эволюции палеофауны и в определенной мере палеофлоры. Недавно были получены данные о том, что некоторые виды радиолярий чувствительны к магнитным инверсиям. По палеоданным около дюжины видов радиолярий, некогда обитавших в различных акваториях в площади современного Тихого океана, вымерли геологически мгновенно и повсеместно примерно 2,5 млн. лет назад на рубеже так называемой палеомагнитной эпохи Гаусс.
Как утверждает Е.П. Дубинин (1997), современное развитие эволюционной палеонтологии имеет существенную возможность для определенных экологических реконструкций. В мезозое, как известно, высочайшего расцвета достигли рептилии, пресмыкающиеся, которых за 200 млн. лет сформировалось примерно 20 отрядов. В кайнозое же максимальный расцвет пришелся на млекопитающих, причем примерно за 70 млн. лет образовалось уже 30 их отрядов. Это объясняется сравнением условий развития рептилий и млекопитающих. Бурное развитие рептилий совпадает с образованием Пангеи, с регрессией океана и мягким климатом. В целом в этих условиях во всех пределах суши существовала единая экологическая провинция и практически не было экологически изолированных или полуизолированных провинций. А это, как известно, не располагает к формированию разнообразия среды обитания рептилий, что подтверждается фактом того, что в самом конце палеозоя, когда сформировались более или менее изолированные материковые фрагменты, именно в это время из 20 отрядов сформировалось более чем 10 новых отрядов рептилий.
В начале кайнозоя, когда Пангея практически разрушилась, на отдельных осколках материков существовало некоторое количество отрядов примитивных млекопитающих. Именно "разбегание" материков в позднем мелу тогда, когда млекопитающие приобрели тенденцию к расцвету, привело к определенной изоляции отдельных континентов и при морской трансгрессии и к формированию почти полностью отдельно развивающихся экологических провинций. А это привело к возникновению множества отрядов млекопитающих. В позднем же кайнозое несколько материковых фрагментов соединилось, что выразилось в вымирании 13 отрядов сухопутных млекопитающих. При этом в конкурентной борьбе выжили те, которые в палеогене и начале неогена развивались в "пограничных" ("опушечных") экологических провинциях.
Намеченная выше линия эволюции биосферы с момента начала цефализации (от греч. кефа! - голова), когда организмы приобрели способность реагировать на окружающий мир, т. е. чувствовать, стала резко менять свою кривизну, начала интенсивно расти. Именно в это время к концу мезозоя свершилось величайшее открытие в жизненных формах - изобретение амниотического яйца. Яйцо разительно отличается от икринок и, как известно, содержит в себе всю необходимую для дальнейшего развития зародыша микросреду, включая даже запасы воды. Благодаря этим особенностям амни-отические яйца могли быть оставлены в самых недоступных для хищников местах, и рептилии получили уникальную возможность страхования продолжения жизни. В тех глобальных экологических условиях - соотношениях и конфигурации океана и материков, химизме атмосферы и гидросферы, геомагнитной обстановке и космических факторах - это в целом и предопределило расцвет рептилий.
Освоение суши в последующие геологические эпохи растениями, совершенствование которых тоже было весьма активным, сформировало на континентах наиболее
111
сложную из всех известных экосистем - лес. Высшая форма эукариотов, адаптировавшаяся к лесу в кайнозое в результате изобретения плаценты в форме млекопитающих, получила в новой экологической обстановке преимущество как теплокровных над холоднокровными рептилиями. Среди млекопитающих именно в лесу появился новый лидер эволюции - приматы. Это довольно слабое, весьма скромных размеров животное без средств защиты и нападения в виде когтей, бивней, панциря и т. п., имело удивительную способность к совершенствованию. Они достаточно быстро приобрели хватательные способности и стереоскопическое зрение, что позволило получить преимущества в защите и нападении, и в частности, за счет уникальной способности к совершенствованию сложнейших прыжков.
Приматы быстро освоили наиболее продуктивную верхнюю зону леса, сохранив возможность посещения поверхности земли и совершения путешествий между лесными массивами. Появление среди приматов человека, точнее его прямых предков примерно 2 млн. лет тому назад (по некоторым данным, например, публикациям А. Лики гораздо раньше около 10 млн. лет тому назад) в геологическом (палеонтологическом) смысле событие незначительное. Но в информационном смысле, показателя темпов эволюции, появление рода Homo sapiens предшествовало образованию человеческого общества как значительного фактора во взаимодействиях с биосферой Земли, что, как показывает современный опыт, имеет принципиальное значение.
Разные подходы к оценке факторов, создавших человека, обусловливались в конечном счете уровнем научного знания и его интерпретациями. Среди значительного числа специалистов определенное время господствовало представление о том, что "труд сделал человека из обезьяны" (Ф. Энгельс. "Диалектика природы" 1882, 1925 г.). Однако еще задолго до появления прямых предков человека приматы обладали способностью использовать орудия труда (палкой - эпизодически еще 10-15 млн. лет тому назад), камнем - 2-3 млн. лет тому назад. Во многом это было следствием просто изменений в климате - леса превратились в саванны и для защиты и нападения способные к подражанию приматы "вооружились когтями из палок" и увеличили свою силу за счет веса камня в руке-лапе. Австралопитеки, собирая пищу в виде кореньев, плодов, падали и мелких живых организмов, использовали эти орудия труда опять же эпизодически, фрагментарно передавая этот навык в наследственной памяти, не формируя его адаптационной сущности. В.А. Зубаков пишет, что австралопитеки в виде небольших стад в 10-20 особей существовали в саванне и, невзирая на эпизодическое пользование орудиями труда, подчинялись правилам биологического доминирования. Стадо возглавлялось сильнейшим самцом, что приводило к тому, что при его доминировании потомство создавалось именно этим самцом, потому что при эструсе - время половой возбудимости - (у самок австралопитеков он не превышал, как и у обезьян, 5-6 дн) ни один другой самец не мог противиться вожаку, но потомство не всегда было лучшим.
Важнейшим следующим этапом в развитии, точнее, в создании собственно человека, была сексуальная революция, которая отделила собственно человека от животных. Это утверждение В.А. Зубакова мы считаем биологически и экологически обоснованным. Именно этот этап резко изменил течение эволюции, произошел новый качественный скачок в развитии приматов - пралюдей. Сексуальная революция свершилась внутри стада австралопитеков, возможно только в одном случайном стаде, когда у одной из самок опять же случайно в результате мутаций эструс стал постоянным. Считаем весьма важным процитировать слова В.А. Зубакова (1997, с. 177): "Появление первой женщины (Евы - Прародительницы, прим. автора), т. е. самки с постоянной половой возбудимостью, в корне изменило взаимоотношения самцов и открыло путь к количественному росту стада, а значит, и числа добытчиков и охранников. Мяса стало достаточно для всех, в том числе и для стариков, которые заготавливали и обрабатывали камни для охоты и при этом могли обучать детей". В целом сформировались условия для замены биологического доминирования в таком стаде на социальные. Естественно, что единственный случай такой мутации не мог в корне изменить все сложившиеся взаимодействия во всех стадах австралопитеков, но возможная наследственная передача способности к постоянному эструсу при большом количестве потомства и вероятная предрасположенность
112
самок австралопитеков к такой мутации в других стадах в целом создали предпосылки к социальной революции и сформировали Homo sapiens.
Сложившиеся новые условия в существовании предков человека значительно стимулировали развитие мозга за счет возможностей передачи информации от ее носителей, имеющих жизненный опыт. Одним из дополнительных факторов формирования социальных отношений стала необходимость поддержания огня. Пралюди весьма длительный период времени не умели разводить огонь, но поддерживали его десятками лет, а для этого нужно было иметь вполне определенные уже отнюдь не биологические взаимоотношения внутри стада. Как пишет В.А. Зубаков, костер был исключительной точкой, вокруг которой начала зарождаться система обмена культурной информацией - возникла знаковая система и, наконец, язык и его носитель - Учитель. Собственно, это и стало началом Социума (общества людей).
Дальнейшее развитие человека как одного из представителей биосферы довольно значительный период времени находилось в соответствии с окружающей средой, люди чувствовали себя частью природы и находились с ней в реальном гомеостазе. Собственно в это время человек сформировал первые мифологические представления об окружающем мире, именно в это время человек "обожествил" природу и одновременно "оживил" и "очеловечил" ее. Это продолжалось практически весь палеолит и только в неолите возникли серьезные конфликты с природной средой, что выразилось в квазиглобальном экологическом кризисе. Человек к этому времени освоил навыки загонной охоты с огнем, не нанося серьезного урона животным до тех пор, пока не возникли изменения глобального уровня, не наступила пора межледниковья с резким потеплением, что нанесло серьезный удар по предмету охоты человека, по мамонтовой популяции и по крупным копытным. Резкое снижение запасов пищи поставило в свою очередь популяцию человека на грань почти полного исчезновения. Выход из кризисной ситуации в неолите был найден в переходе человека к земледелию и скотоводству. С этого времени и начинается эпоха развития человеческого общества, изучение которой является предметом исторической науки.
По мере формирования структуры планеты происходило перемещение химических элементов и их соединений: легкие из них, такие, как Н2, СО2, СН4 и другие, оставались на поверхности, а тяжелые перемещались вглубь. Атмосфера состояла из свободного водорода и его соединений (СН4, NH3, HCN, Н2О) и носила восстановительный характер. Академик А.И. Опарин считал, что именно это было главной предпосылкой возникновения органических молекул небиологическим путем. Еще в XIX в. было установлено, что органические молекулы можно синтезировать в лабораторных условиях (Ф. Велер), но большинство ученых все равно считали, что эти соединения возникают исключительно в живых организмах (отсюда собственно и их название-"органические"). Но астрономы обнаружили метан в составе атмосферы Сатурна, Юпитера и в ряде туманностей; принципиально было доказано, что простейшие углеводороды могут образовываться в космическом пространстве, а это означает, что они могли входить в состав первичной атмосферы Земли. Вместе с другими компонентами - аммиаком, водородом, синильной кислотой, парами воды - они подвергались воздействию различных источников энергии: жесткому, близкому к рентгеновскому и ультрафиолетовому излучению от Солнца, высокой температуре при вулканических извержениях и в зонах грозовых разрядов. В результате создавались условия для протекания все более сложных реакций. Возникали молекулы Сахаров, аминокислот, азотистых оснований, органических кислот и т. п. С. Миллер в 1953 г. экспериментально подтвердил возможность таких превращений. В своей знаменитой "колбе Миллера" он, пропустив электрический разряд через смесь Н2, Н2О, СН4 и NH3, получил набор из нескольких аминокислот и органических кислот. Далее было установлено, что абиогенным путем в отсутствие кислорода синтезируются
113

Рис. 32. Схема, отражающая симбиотическую гипотезу возникновения эукариотических клеток
многие простые органические соединения, входящие в состав биологических полимеров - белков, нуклеиновых кислот и полисахаридов. Органические соединения были обнаружены в космическом пространстве, в частности цианистый водород (HCN), формальдегид, муравьиная кислота, этиловый спирт. В некоторых метеоритах были найдены жирные кислоты, сахара, аминокислоты.
В условиях первичной атмосферы и гидросферы Земли происходила конденсация простейших органических соединений, в результате чего образовывались первичные полимеры-полипептиды и полинуклеотиды. Надо заметить, что для дальнейшего образования более сложных органических веществ требуются гораздо менее жесткие условия, чем для образования простых молекул. По С.Г. Мамонтову (1999), синтез аминокислот из смеси газов, входивших в состав атмосферы древней Земли, происходит при температуре 1000° С, а превращение их в полипептиды - всего лишь при 160° С (рис. 32).
Вполне утвердительно можно сказать, что образование разнообразных органических соединений из неорганических веществ в тех условиях было закономерным процессом химической эволюции.
Первичный океан, по-видимому, содержал в растворенном виде различные органические и неорганические молекулы, попадающие в него из атмосферы и вымываемые из поверхностных слоев Земли. Концентрация органических соединений постоянно увеличивалась, и в конце концов вода океана стала "бульоном" из белковоподобных веществ - пептидов, а также нуклеиновых кислот и других органических соединений. Молекулы различных веществ могут соединяться, образуя многомолекулярные комплексы - коацерваты (рис. 33,34). В первичном океане коарцеваты, или коацерватные капли, обладали способностью поглощать различные вещества, растворенные в его водах. В
114
Рис. 33. Образование водной оболочки вокруг крупных молекул и формирование коацерватов
результате этого внутреннее строение коарцевата претерпевало изменения, что приводило или к его распаду, или к накоплению веществ, т. е. к росту и изменению химического состава. Это, в свою очередь, вело к повышению устойчивости коацерватной капли в непрерывно изменяющихся условиях среды. Нужно сказать, что судьба капли: распад или рост, определялась преобладанием процесса. Академик А.И. Опарин утверждал, что преобладание указанного процесса обусловливалось некоторым отбором коацерватных капель па устойчивости к данным конкретным условиям. Выросшая до определенных размеров "материнская" коацерватная капля могла распадаться на дочерние. Эти образовавшиеся капли начинали расти и повторяли после определенного этапа распад на дочерние. Большое значение имело то, что материнские и дочерние капли были близки по составу, а отличные от них подпадали под процесс распада. Естественно, что продолжали существовать только те коацерватные капли, которые, взаимодействуя через элементарные формы обмена со средой, сохраняли относительное постоянство состава. В дальнейшем, как полагает С.Г. Мамонтов (1999), они приобрели способность поглощать из окружающей среды лишь те вещества, которые обеспечивали их устойчивость; вторым приобретенным свойством для них стала возможность выделять наружу продукты обмена. Такие процессы постепенно создали различия между химическим составом капли и окружающей средой. Собственно
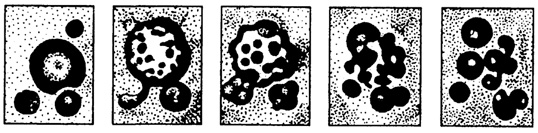
Рис. 34. Коацерватные капли, полученные в экспериментальных условиях
результатом химической эволюции явилось сохранение только тех коацерватных капель, которые при распаде материнской капли на дочерние не утрачивали своих структурных особенностей и состава, т. е. приобретали важнейшую способность жизни - самовоспроизведение.
Следует сказать, что это наиглавнейшее свойство возникло совместно с расположенностью к синтезу органических веществ внутри коацерватных капель. Важнейшими составными частями этих образований были полипептиды и полинуклеотиды. Между синтезом и свойством самовоспроизведения возникла неразрывная связь. Химические превращения органических соединений привели к образованию полипептидов, обладающих каталитической активностью, т. е. способностью значительно ускорять течение химических реакций.
Полинуклеотиды, как известно, способны осуществлять неферментативный синтез дочерних нуклеотидных связей, т. е. связываться по принципу дополнения или комплементарности.
Следующим этапом небиологической эволюции органических соединений стало объединение способности полинуклеотидов к самовоспроизведению с каталитическими возможностями полипептидов, так как удвоение молекул ДНК (дезоксирибонуклеиновой кислоты) эффективнее осуществляется с помощью белков, обладающих способностью к катализу. При этом стабильность устойчивых комбинаций аминокислот обеспечивается исключительно сохранением информации о них в нуклеиновых кислотах.
115
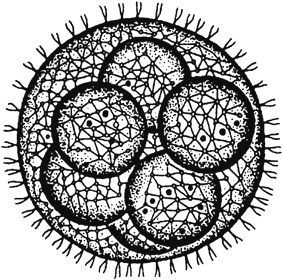
Рис. 35. Современный свободно движущийся представитель колониальных простейших - вольвокс
Связь белковых молекул и нуклеиновых кислот привела к возникновению генетического кода. Последний представляет собой такую организацию молекул ДНК, в которой последовательность нуклеотидов стала служить информацией для построения конкретной последовательности аминокислот в белках.
С.Г. Мамонтов и В.Б. Захаров (1999) пишут, что дальнейшее усложнение обмена веществ у предбиологических структур могло происходить только в условиях пространственного разделения различных синтетических и энергетических процессов внутри коацервата, а также более прочной изоляции внутренней среды от внешних воздействий по сравнению с той, которую могла осуществить водная оболочка. Эту изоляцию стала осуществлять мембрана, которая предположительно возникла следующим образом: вокруг коацерватов, богатых органическими соединениями, возникли оболочки (концентрические слои) жиров, или липидов, отделившие коацерват от окружающей водной среды и преобразовавшиеся в ходе дальнейшей эволюции собственно в мембрану. Появление биологической мембраны, отделившей содержимое коацервата от окружающей среды и обладающей способностью к избирательной проницаемости, предопределило направление дальнейшей химической эволюции по пути развития все более совершенных саморегулирующихся систем, вплоть до возникновения первых примитивно устроенных клеток (рис. 35).
С момента появления клеточных структур химическая эволюция органических соединений сменилась биологической эволюцией.
По предположению акад. Б.С. Соколова жизнь на Земле существует 4 млрд. 250 млн. лет (!). Именно здесь прослеживается граница между отсутствием и наличием жизни на нашей планете. По геологическим масштабам жизнь возникла почти мгновенно - всего за 250 млн. лет после образования Земли практически одновременно с формированием Мирового океана. Современные научные данные показывают, что первые клеточные образования появились еще через 1 млрд. лет, следы этих первых организмов обнаружены в породах, имеющих возраст 3,1 млрд. лет. Размеры этих первых обитателей очень малы (рис. 36).
Очень многие факторы среды обеспечивали появление жизненных форм на Земле, а в дальнейшем поддерживали и

Рис. 36. Остатки первых микроорганизмов в древних горных породах:
а -
в - возраст 1,9 млрд. лет;
г, д - возраст 3,1 млрд. лет
116
определяли функционирование жизненных форм в условиях нашей планеты. Например, Дж. Бернал установил, что глинистые отложения на дне Мирового океана сослужили роль аккумулятора органических соединений абиогенного происхождения. Ряд специалистов утверждают, что на ранних стадиях существования Земля проходила в космосе через пылевые облака и захватила вместе с космической пылью большое количество органических молекул, образовавшихся в космическом пространстве. Количество этих молекул, как считают некоторые ученые, соизмеримо с биомассой современной Земли.
Архейская и протерозойская эры. Докембрий - 4,6 млрд. лет назад. Земная кора и атмосфера находятся в процессе формирования. В течение докембрия древние горные породы подвергались складкообразованию, сдвигам, метаморфизму и эрозии. В раннем докембрии Земля была достаточно горячей. С тех пор она постепенно охлаждается. Первая известная нам эпоха оледенения имела место около 2,3 млрд. лет тому назад, позднее в докембрии было еще два оледенения. Величайшее же оледенение в истории планеты произошло между 1 млрд. и 600 млн. лет тому назад.
Около 3,5 млрд. лет назад появились первые одноклеточные организмы. Примерно 2,8 млрд. лет назад, или немногим ранее, возникли первые "фотосинтезаторы" (стромалиты), и содержание кислорода стало медленно повышаться. Первые многоклеточные организмы появились около 1,4 млрд. лет назад, а первые клетки с центральным ядром - примерно около 1,2 млрд. лет назад. В позднем докембрии возникли плоские и сегментированные черви, медузы и иглокожие. Растительность отсутствует.
Палеозойская эра ("эра древней жизни"). Кембрий - 570 млн. лет назад. Сверхматерик Гондвана занимает экватор и приэкваториальные части планеты. Наряду с Гондваной было еще четыре материка меньших размеров, примерно соответствующих по площади современным Европе, Сибири, Китаю и Северной Америке. В мелких тропических водах формируются обширные строматолитовые рифы. На суше происходила интенсивная эрозия и большое количество осадков сносилось в моря. Содержание кислорода постепенно повышалось. Ближе к окончанию периода началось оледенение, приведшее к понижению уровня воды в морях.
В процессе грандиозного эволюционного взрыва возникло большое число типов животных, многие из которых существуют и сейчас, включая микроскопических ежей, морских лилий и различных червей. В тропиках археоциаты возводили громадные рифовые сооружения. Появились первые твердопокровные животные; в морях господствовали трилобиты и брахиоподы. Возникли первые хордовые. Несколько позднее появились головоногие моллюски и примитивные рыбообразные. Из растительности следует отметить наличие примитивных морских водорослей.
Ордовик - 500 млн. лет назад. Гондвана находится в южном полушарии, а остальные материки несколько переместились к экватору. "ПраЕвропа" и "ПраСеверная Америка" медленно отходят друг от друга и одновременно океан Япетус расширяется. На протяжении всего периода массивы суши смещались все дальше и дальше к югу. Старые ледниковые покровы кембрия растаяли, в связи с чем произошло поднятие уровня моря. Большая часть суши сконцентрировалась в теплых широтах. В конце периода началось новое оледенение.
Резко увеличивается численность животных - фильтратов, в том числе мшанок ("морских циновок"), морских лилий, плеченогих двустворчатых моллюсков и граптолитов (для последних это было время расцвета). Археоциаты уже вымерли, но строительство рифов продолжалось первыми кораллами и строматопорондеями. Увеличилось число наутилоидей и бесчелюстных панцирных рыб.
Развились различные виды водорослей, а в позднем ордовике появились первые настоящие наземные растения.
Силур - 438 млн. лет назад. Гондвана надвинулась на Южный полюс. Океан Япетус постепенно начал уменьшаться в размерах и массивы суши "ПраСеверная Америка" и
117
"ПраГренландия" стали сближаться. В конечном счете они столкнулись, образовав гигантский материк Лавразию. Это был период активной вулканической деятельности и интенсивного горообразования. Начался этот процесс с эпохи оледенения, когда же льды растаяли, то уровень моря повысился и климат стал мягким.
Ругозы ведут активное строительство рифов. Численность граптолитов снижается. В морях процветают наутилоиды, брахиоподы, трилобиты и иглокожие. Вода в морях обладает невысокой соленостью и в ней обитают ракоскорпионы (эвриптериды). Изобилие рыб как в пресной, так и в соленой воде. Появились первые челюстные рыбы - акантоды. Скорпионы, многоножки и, как предполагают, эвриптериды постепенно выходили на сушу.
Растения заселили берега водоемов. Преобладают примитивные псилопсидные растения.
Девон - 408 млн. лет назад. В южном полушарии раскинулась Гондвана. В тропиках продолжается формирование Лавразии. Происходит интенсивная эрозия недавно образовавшихся гор, в результате чего образуются мощные отложения красного песчаника; речные дельты в связи с большим количеством поступающих наносов заболачиваются. Уровень моря к концу периода поднимается. Средние температуры воздуха возрастают, но в целом он становится более резким, с чередованием периодов ливневых дождей и сильной засухи. Обширные площади на материках стали безводными.
Быстрая эволюция рыб, включая акул и скатов, кистеперых и лучеперых рыб. Увеличилось число аммонитов. В морях развились гигантские хищники - эвриптериды длиной до 2 м. В позднем девоне многие группы древних рыб, а также кораллов, плеченогих и аммонитов вымерли. Суша подверглась нашествию множества членистоногих, в том числе клещей, пауков и примитивных бескрылых насекомых. В позднем девоне появились первые земноводные.
Растения сумели отодвинуться от кромки воды, и постепенно обширные районы суши заросли густыми первобытными лесами. Возросло число разнообразных сосудистых растений. Появились споровые ликофиты (плауны) и хвощи, некоторые развились до размеров настоящих деревьев высотой почти 40 м.
Карбон (каменноугольный) - 360 млн. лет тому назад. Гондвана и Лавразия постепенно сближались, образовывая новые горные цепи. В раннем карбоне на обширных пространствах образовались мелкие прибрежные моря и болотистые низменности; на большой части суши установился почти тропический климат. Наличие громадных лесов с пышной растительностью способствовало повышению содержания кислорода в атмосфере. Ближе к окончанию периода началось похолодание, и на Земле произошло два крупных оледенения.
В морях появились новые разновидности аммонитов, возросла численность брахиоподов. Ругозы, граптолиты, трилобиты, а также некоторые мшанки, морские лилии вымерли. Это было время расцвета земноводных, а также насекомых-кузнечиков, тараканов, чешуйниц, термитов, жуков и гигантских стрекоз. В позднем карбоне появились первые рептилии.
Дельты рек и берега обширных болот поросли густыми лесами из гигантских плаунов, хвощей, древовидных папоротников и семенных растений высотой до 45 м. Неразложившиеся останки этой растительности со временем превратились в залежи каменного угля.
Пермь - 286 млн. лет назад. Гондвана и Лавразия еще больше сблизились. "Пра-Индостан" и Азия столкнулись, и возник гигантский сверхматерик Пангея. Это столкновение также породило новые горные цепи. Пангея начала перемещаться к северу. Этот период начался с оледенения, которое вызвало понижение уровня моря. По мере движения Гондваны к северу земная кора прогрелась и льды постепенно растаяли. В Лавразии стало очень жарко и сухо, по ней распространились обширные пустыни.
Бурно эволюционировали двустворчатые моллюски. В морях в изобилии водились аммониты. Главными строителями рифов постепенно становятся современные кораллы. В ранней перми в пресных водоемах господствовали земноводные. Появились и водные рептилии, в том числе мезозавры. В ходе великого вымирания в конце периода полностью
118
исчезло свыше 50% животных семейств, включая многих земноводных, аммонитов и трилобитов. На суше рептилии взяли верх над земноводными.
На южных массивах суши распространились леса крупных семенных папоротников - глоссоптерисов. Появились первые хвойные, быстро заселившие внутриматериковые области и высокогорья.
Мезозойская эра ("эра средней жизни"). Триас - 248 млн. лет назад. Гондвана вновь начала разделяться, стал образовываться Атлантический океан. Уровень моря по всему миру был очень низок. Климат, почти повсеместно теплый, постепенно становился более сухим, и во внутриматериковых областях сформировались обширные пустыни. Мелкие моря и озера интенсивно испарялись, из-за чего вода в них постепенно становилась все более соленой.
Динозавры и прочие рептилии стали доминирующей группой наземных животных. Появились первые лягушки, а чуть позже сухопутные и морские черепахи и крокодилы. Возникли также и первые млекопитающие, возросло разнообразие моллюсков. Образовались новые типы кораллов, креветок и омаров. К концу периода вымерли почти все аммониты. В океанах утвердились морские рептилии, такие, как ихтиозавры, а птерозавры начали осваивать воздушную среду.
Возросло разнообразие голосеменных растений, образовавших обширные леса саговников, араукарий, гинкго и хвойных деревьев. В нижней части леса сформировались ковры из плаунов и хвощей, а также пальмовидных беннеттитов.
Юра - 230 млн. лет назад. Пангея продолжала раскалываться, и море затопило большую часть суши. Происходило интенсивное горообразование. В начале периода климат был повсеместно теплым и сухим, затем стал более влажным.
Увеличилось количество и стали более разнообразными морские черепахи и крокодилы, появились новые виды плезиозавров и ихтиозавров. На суше процветали насекомые, включая предшественников современных муравьев, пчел, уховерток, мух и ос. Появилась и первая птица - археоптерикс. Господствовали динозавры, эволюционировавшие во множество форм - от зауроподов до более мелких быстроногих хищников.
Климат стал более влажным, и вся суша поросла обильной растительностью. В лесах появились предшественники нынешних кипарисов, сосен и мамонтовых деревьев.
Мел - 114 млн. лет назад. С удалением материков друг от друга Атлантический океан, разделяющий Африку и Южную Америку, расширился до весьма значительных размеров. Африка, Индостан и Австралия, находившиеся на экваторе и южнее него, приобрели направление раздвижения. Море затопило обширные участки суши. Останки твердопокровных планктонных организмов образовали на океанском дне огромные толщи отложений писчего мела и мелоподобных пород. Поначалу климат был теплым и важным, однако затем наступило весьма заметное похолодание.
В морях возросло количество белемнитов. В океанах господствовали гигантские морские черепахи и хищные морские рептилии. На суше появились змеи, кроме того, возникли новые разновидности динозавров, а также насекомых, таких, как мотыльки и бабочки. В конце с периода произошло гигантское массовое вымирание: исчезли аммониты, ихтиозавры и многие другие морские животные, а на суше вымерли все динозавры и птерозавры.
Появились первые цветковые растения, которые создали "взаимообслуживающие" связи с насекомыми, которые переносили пыльцу и частично ею питались. Эти растения весьма активно распространялись по всему пространству суши.
Кайнозойская эра ("эра новой жизни"). Палеоген. Палеоцен - 65 млн. лет назад. Южные материки продолжали раскалываться. Южная Америка оказалась полностью отрезанной от остальных участков суши. Африка, Индостан и Австралия разошлись друг от друга на значительные расстояния, при этом Австралия заняла позицию вблизи Антарктиды. Обнажились новые участки суши и произошло дальнейшее понижение уровня моря.
На суше началось время активного развития млекопитающих. Появились грызуны и насекомоядные, "планирующие" млекопитающие и ранние приматоподобные. В морях
119
на смену морским рептилиям пришли новые виды хищных костных рыб и акул. Возникли новые разновидности двустворчатых моллюсков и фораминифер.
Продолжали распространяться все новые виды цветковых растений и опылявших их насекомых.
Эоцен - 55 млн. лет назад. Индостан придвинулся к Азии. Антарктида и Австралия в начале периода располагались рядом, но в дальнейшем начали расходиться. Северная Америка и Европа также разделились, при этом возникли новые горные цепи.Море затопило часть суши. Климат повсеместно был теплым. На суше появились летучие мыши, лемуры, долгопяты; предки нынешних слонов, лошадей, коров, свиней, носорогов и оленей, а также прочие крупные травоядные. Другие млекопитающие типа китов и сирен вернулись в водную среду. Увеличилось число видов пресноводных костных рыб. Эволюционировали и другие группы животных, в том числе муравьи и пчелы, скворцы и пингвины, гигантские нелетающие птицы, кроты, верблюды, кролики и полевки, кошки, собаки и медведи и др.
Во многих частях света произрастали леса с пышной растительностью, в умеренных широтах росли пальмы.
Олигоцен - 38 млн. лет назад. Индостан пересек экватор, Австралия полностью оторвалась и отошла от Антарктиды. Климат стал прохладнее, над Южным полюсом сформировался огромный ледяной покров, что привело к понижению уровня моря.
С распространением степей начался бурный расцвет травоядных млекопитающих. Среди них возникли новые виды кроликов, зайцев, гигантских ленивцев, носорогов и прочих копытных. Появились первые жвачные.
Тропические леса уменьшились в размерах и начали уступать место лесам умеренного пояса, появились и обширные пространства со степной - травянистой - растительностью. Быстро распространились новые виды трав, к которым приспосабливались новые виды травоядных животных.
Неоген. Миоцен-25 млн. лет назад. Африка столкнулась с Европой и Азией, образовав Альпы. Индостан "врезался" в Азию и воздвиг Гималаи. По мере наползания материковых плит друг на друга начали формироваться Альпы и Скалистые горы. Ледниковый покров в южном полушарии закрыл всю Антарктиду, что привело к дальнейшему охлаждению климата. Млекопитающие мигрировали с материка на материк по новообразовавшимся сухопутным мостам, что резко ускорило эволюционные процессы. Слоны из Африки перебрались в Евразию, а в обратном направлении началась миграция буйволов, жирафов, свиней, кошек. Появились саблезубые кошки и обезьяны, в том числе человекообразные. В отрезанной от других материков Австралии продолжали развиваться однопроходные и сумчатые.
Внутриматериковые области становились все холоднее и засушливее и в них все больше распространялись степи.
Плиоцен - 5 млн. лет назад. Материки почти достигли их современного положения. Громадные ледниковые покровы распространились по всему северному полушарию; такое же оледенение было и в Антарктиде, на юге Южной Америки. Климат стал еще более холодным.
Травоядные копытные млекопитающие продолжали бурно размножаться и эволюционировать. Ближе к концу периода сухопутный мост связал Южную и Северную Америку, что привело к грандиозному "обмену" животными между двумя материками. Полагают, что обострившаяся межвидовая конкуренция вызывала вымирание многих древних животных. В Австралию проникли крысы, а в Африке появились первые человекоподобные существа. По мере охлаждения климата леса заменились степями.
Четвертичный. Плейстоцен - 2 млн. лет назад. Это была эпоха великого оледенения с чередованием периодов похолодания и потепления и колебаниями уровня моря. Эта ледниковая эпоха продолжается и по сей день.
Некоторые животные сумели адаптироваться к усилившимся холодам, обзаведясь густой шерстью, к примеру шерстистые мамонты и носороги. Из хищников наиболее распространены саблезубые кошки, пещерные львы. Это было время расцвета гигантских сумчатых в Австралии и громадных нелетающих птиц типа моа или эпиорнисов,
120
обитавших во многих районах южного полушария. Появились (?!) первые люди; многие крупные млекопитающие начали постепенно исчезать.
С полюсов постепенно наползали льды и хвойные леса уступали место тундре. Дальше от края ледника лиственные леса стали заменяться хвойными. В более теплых областях земного шара раскинулись обширные степи.
Голоцен-0,01 млн. лет назад. В течение всего голоцена материки занимают практически современные места, климат очень похож на нынешний, только каждые несколько тысячелетий становясь то теплее, то холоднее. В настоящее время имеет место один из периодов потепления. По мере уменьшения ледниковых покровов уровень моря поднимался.
В настоящий период многие виды животных вымерли в основном из-за усиленной охоты человека на них; при перемещении животных в другие места Земли возникла новая конкуренция. Человеческая цивилизация постепенно развилась до современного состояния.
С возникновением земледелия крестьяне уничтожали все больше дикорастущих растений, возникли агроценозы, сформировались современные растительные сообщества.