Глава 14
Естественное равновесие и эволюция
экосистем
Рассмотрение понятий о равновесии и неравновесии систем с синергетических позиций позволило выявить основные отличия между этими двумя возможными их состояниями систем (по А.А. Горелову, 1998):
- система способна к реакциям на внешние условия;
- поведение системы обусловлено ее предысторией, но зависит от начальных условий и является случайным;
- приток энергии создает упорядоченность в системе, что влечет за собой уменьшение ее энтропии;
- система ведет себя как единое целое, как если бы она была вместилищем дальнодействующих сил (это отвечает некоторым физическим гипотезам); несмотря на то, что силы молекулярного взаимодействия представляют собой короткодействующие (т. е. на расстоянии до 10-8 см), система структурируется так, как если бы каждая молекула была "информирована" о состоянии системы в целом.
В целом можно выделить области равновесности и неравновесности для систем, при которых существенно различается поведение системы (табл. 15).
Любая система при отсутствии какого-либо воздействия, т. е. будучи предоставленной самой себе и при отсутствии поступления энергии (а по последним исследованиям и информации) извне, стремится к наиболее вероятному состоянию равновесия, достигнутому при энтропии, равной нулю. Примером такой равновесной структуры служит кристалл минерала, "выросшего" в идеальных условиях. К данному равновесному состоянию в соответствии со вторым началом
210
термодинамики стремятся и приходят все закрытые системы, т. е. системы, не получающие энергии и информации извне. Системы обратного типа называют открытыми.
Таблица 15. Характеристика состояния систем
Неравновесная область
Равновесная область
- 1. Система "адаптируется" к внешним
условиям, изменяя свою структуру.
- 2. Множественность стационарных
состояний.
- 3. Чувствительность к флукгуациям (наибольшие
влияния приводят к большим
последствиям, внутренние флуктуации
становятся большими).
- 4. Все части системы действуют
согласованно.
- 5. Фундаментальная неопределенность.
- 1. Для перехода от одной структуры
к другой требуются сильные возмущения
или изменение граничных условий.
- 2. Одно стационарное состояние.
- 3. Нечувствительность к флукгуациям.
- 4. Молекулы, например, ведут себя
независимо друг от друга.
- 5. Поведение системы определяют
линейные зависимости.
Изучение неравновесных состояний для различных систем позволяет выявить общие закономерности эволюции от хаоса к упорядоченности.
Особенности эволюции. Еще в Древней Греции было известно различие между "хаосом" и "космосом". Современные достижения в синергетике позволяют назвать хаотическими все системы, которые приводят к несводимому представлению, если их оценивать в терминах теории вероятности. Иными словами, эти системы нельзя описать однозначно детерминистично (определенно), т. е. точно предсказать, что будет с системой в будущем, зная состояние системы в настоящем.
Хаотическое поведение непредсказуемо в принципе. Необратимость, вероятность и случайность становятся объективными свойствами хаотических систем на макроуровне, а не только, как было установлено в квантовой механике, на микроуровне.
С синергетической точки зрения эволюция должна отвечать следующим условиям:
- необратимости, выражающейся в нарушении симметрии между прошлым и будущим;
- необходимости наличия понятия "событие";
- способности некоторых событий изменять ход эволюции. Известно, что при любом развитии эволюция создает возможность
- формирования новых структур, для которого необходимо выполнение следующих условий:
- открытость системы;
- нахождение системы на "удалении" от равновесия;
- наличие флуктуации (случайного отклонения).
211
Чем сложнее система, тем более многочисленны флуктуации и их типы. Однако в сложных системах существуют связи между различными ее частями. От соотношения устойчивости, которая обеспечивается взаимосвязью частей, и неустойчивости из-за флуктуации зависит порог устойчивости системы. При переходе через этот порог система попадает в критическое состояние, называемое точкой бифуркации. В ней система становится неустойчивой относительно флуктуации и может перейти к новой области устойчивости. При этом система как бы стоит перед выбором какого-то из нескольких путей эволюции. Наибольшая флуктуация может послужить в этот момент началом эволюции в совершенно новом направлении, который резко изменит поведение системы, что собственно и есть событие.
В точке бифуркции случайность толкает систему на новый путь развития, а после того, как один из возможных вариантов выбран, вновь вступает в силу определенность (детерминизм) вплоть до следующей точки бифуркции. В развитии, в судьбе, системы случайность и необходимость являются взаимно дополняющими условиями.
Большинство известных (и изученных) систем представляют собой открытые системы, которые обмениваются энергией, веществом и информацией с окружающей средой. Как показали исследования, главенствующую роль в природе играют не порядок, стабильность и равновесие, а неустойчивость и неравномерность, т. е. все системы флуктуируют. В некоторой особой точке бифуркации флуктуация становится настолько мощной, что система разрушается и в данный момент принципиально невозможно установить, что произойдет с системой: станет ли она по своему состоянию хаотической или перейдет на новый более дифференцированный и высокий уровень упорядоченности, который называют диссипативной (рассеянной) структурой. Новые структуры называют диссипативными, потому что для их поддержания требуется больше энергии, чем для более простых структур, на смену которым они приходят.
Классические исследования в термодинамике XIX века главным образом были направлены на изучение механического действия теплоты и при этом рассматривались закрытые системы, стремящиеся к состоянию равновесия. В настоящее время синергетика (от "синергия" - совместные действия) и термодинамика изучают открытые системы, как правило, далекие от равновесного состояния. Полученные результаты исследований позволяют выявить побудительные причины эволюции в природе; везде, где происходит образование новых структур, необходим метаболизм - обмен со средой и приток энергии. Можно сделать вывод, что современные данные термодинамического, синергетического изучения систем определяют процесс "творчества" природы.
Принцип равновесия является одной из наиболее важных
212
зависимостей в живой природе. Равновесие существует между видами и смещение его в какую-либо сторону вызывает значительные изменения в экосистемах, к примеру, увеличение количества хищников может привести к исчезновению жертв, а уничтожение хищников увеличивает число жертв, но в данном случае жертвам может не хватить пищи. Естественное равновесие существует и между живыми организмами и окружающей его неживой средой. Великое множество "малых" равновесий поддерживает общее равновесие в природе.
Равновесие в живой природе в отличие от неживой является динамичным, а не статичным, т. е. представляет собой движение вокруг некой точки устойчивости. Если сама точка не изменяется, то такое состояние носит название гомеостаза (гомео - то же, стазис - состояние). Гомеостаз - механизм, посредством которого живой организм, противодействуя внешним воздействиям, поддерживает параметры своей внутренней среды на таком постоянном уровне, который обеспечивает его нормальную жизнедеятельность. В качестве примера можно привести всем нам известное: величину кровяного давления, частоту пульса, температуру тела, т. е. то, что определяет нормальное состояние здоровья. Это состояние регулируется гомеостатическими механизмами, которые функционируют настолько "отлаженно", что мы их практически не замечаем. Если же функционирование механизма нарушено, то возникает не только дискомфорт в организме, но и опасность его гибели.
Природная экосистема (биогеоценоз) устойчиво функционирует при постоянном взаимодействии ее элементов, круговороте веществ, передаче химической, энергетической, генетической и другой энергии и информации по цепям-каналам. Согласно принципу равновесия любая естественная система с проходящим через нее потоком энергии и информации имеет тенденцию к развитию устойчивого состояния. При этом устойчивость экосистем обеспечивается автоматически за счет механизма обратной связи. Обратная связь заключается в использовании получаемых данных от управляемых компонентов экосистем для внесения корректив управляющими компонентами в процесс. Рассмотренные выше взаимоотношения "хищник" - "жертва" в данном контексте можно описать несколько подробнее; так, в водной экосистеме хищные рыбы (щука в пруду) поедают другие виды рыб-жертвы (карась); если численность карася будет увеличиваться - это пример положительной обратной связи; щука, питаясь карасем, снижает его численность-это пример отрицательной обратной связи; при росте числа хищников снижается число жертв, и хищник, испытывая недостаток пищи, также снижает рост своей популяции; в конце концов в рассматриваемом пруду устанавливается динамическое равновесие в численности и щуки, и карася. Постоянно поддерживается равновесие, которое исключало бы исчезновение любого звена трофической цепи (рис. 64).
213
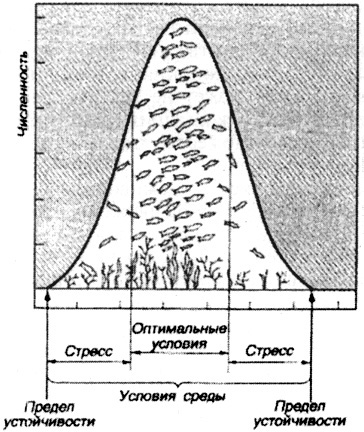
Рис. 64. Наибольшее обилие особей вида в оптимальных условиях
(по Б. Небелу, 1992)
Новые системы обычно подвержены резким колебаниям и менее способны противостоять внешним возмущениям по сравнению со "зрелыми", компоненты которых успели приспособиться друг к другу. Подлинно надежный гомеостатический контроль устанавливается только после периода эволюционного приспособления. Почти всегда, например, существует временная отсрочка в реакциях популяции, которая представляет собой время, необходимое для того, чтобы в неблагоприятных условиях, связанных с перенаселением, начали изменяться рождаемость и смертность в популяции.
Состояние естественного равновесия означает, что экосистема является стабильной и ее отдельные параметры остаются неизменными, несмотря на воздействия, которые система испытывает. Очень важным свойством системы является ее проницаемость - в нее постоянно что-то поступает и постоянно что-то из нее исходит, иными словами, это такое устойчивое состояние экосистемы, при котором поступление вещества, энергии и информации равно их выходу.
В качестве примера действия гомеостатических механизмов рассмотрим динамику популяций. По И.А. Шилову (2000 г. устойчивость популяции, ее относительная самостоятельность, индивидуальные особенности зависят от того, насколько структура и внутренние свойства популяций сохраняют свои приспособительные возможности на фоне изменчивых условий существования. Гомеостаз популяции определяет главное условие - целостность популяции как биологической системы.
Во всех аспектах проявления популяционных функций поддержание их устойчивости имеет абсолютно первостепенное значение. От этого зависят условия функционирования и динамика вышестоящих уровней организации; обеспечение популяционного гомеостаза лежит в основе устойчивого функционирования всего ряда взаимосвязанных систем повышающейся интегрированности (рис. 65). При всем многообразии механизмов гомеостаза популяции можно выделить следующие группы (И.А. Шилов, 2000 г.):
214

Рис. 65. Схема гомеостатического плато
(по Ю. Одуму)
- поддержание адаптивного характера пространственной структуры;
- поддержание генетической структуры;
- регуляция плотности населяющих особей.
Следует заметить, что функции управления гомеостазом популяций изучены к настоящему времени далеко недостаточно, наибольшие результаты получены пока только для животных. Серьезные трудности в изучении этих функций вызывает наличие у высших животных помимо физиологических процессов в реализации гомеостатических функций еще и весьма сложных форм высшей нервной деятельности. Еще более сложными оказываются проблемы изучения гомеостатических функций человеческой популяции, где существуют также недостаточно изученные проявления социума в воздействиях и реакциях.
Отличительные особенности популяционных систем для высших позвоночных животных (как наиболее изученный пример) заключаются в том, что составляющие их элементы-особи (организмы) способны к автономному существованию и не образуют в составе популяции специализированных функциональных систем, наподобие тех, что имеются внутри организма. Важнейшим является то, что все формы взаимодействия популяции со средой и осуществления общепопуляционных функций проявляются опосредованно через физиологические реакции отдельных особей (организмов). Это возможно лишь при определенных формах интеграции деятельности особей, при которых физиологические процессы в отдельных организмах осуществляются в направлении, адаптивном на уровне популяции в целом. Это и есть основа весьма сложных и окончательно еще невыясненных форм внутрипопуляционных отношений, определяющих общий тип и конкретный характер пространственной структуры, уровень и динамику
215
плотности, численности популяции, генетическую структуру и другие свойства популяции и приводящие популяцию в соответствие с условиями среды обитания.
Реализация гомеостатических функций популяции базируется на согласованных действиях составляющих ее особей. Эта согласованность обеспечивается непрерывным потоком информации о состоянии внешней среды и самой популяции. Любой и каждый представитель популяции (особь) одновременно является и источником, и реципиентом ("приемником") информации. Эта информация позволяет особи регистрировать изменения в факторах среды, а главное, инициировать приспособительные реакции от составляющих популяцию особей. Важным моментом является то, что адаптивные ответы на уровне особи интегрируются и выражаются в адаптациях уже на уровне популяции.
Процесс формирования адаптивной реакции на популяционном уровне чрезвычайно сложен и определяется прежде всего разнокачественностью особей по основным эколого-физиологическим свойствам. Это "неодинаковость" каждой из особей популяции и их группировок служит источником неодинаковой информации, так как отдельная особь или даже их группа по-разному реагирует даже на одни и те же факторы среды и поэтому общий "ответ" популяции никогда не является простой суммой ответов отдельных особей. Морфологическая структура популяции (ее "пространственность") обеспечивает оптимизацию процесса адаптивных реакций, но не является непосредственным их "носителем"; физиология и поведение изменяются на уровне отдельных организмов как ответ на полученную информацию. Вследствие этого в отличие от организма - морфологически структурированной системы - популяция может рассматриваться как система информационно-структурированная.
Итак, популяция стабильна, если сохраняет прежде всего свою пространственную структуру. Стремление к восстановлению параметров популяции, способствующей состоянию равновесия, как раз и осуществляется за счет вышеописанных механизмов авторегуляции, которая в конечном счете есть функция следующего уровня организации - экосистемы, частью которой популяция собственно и является.
Выявлено два главных механизма стабилизации плотности популяции при ее величинах, лежащих ниже уровня насыщения: 1) территориальное поведение в виде внутривидовой конкуренции; 2) групповое поведение, выражающееся, например, в "половом доминировании", "порядке клевания" и т. п. В определенной мере эти механизмы проявляются и в человеческом сообществе.
Регулирование популяции может быть физическим и биологическим. Флуктуации численности происходят под влиянием внешних климатических, эдафогенных и других внутренних факторов. Факторы, влияние которых находится в прямой зависимости от плотности
216
популяции, предотвращают перенаселение и способствуют установлению устойчивого равновесия. Главным образом это биотические (конкуренция, паразитизм, патогенное влияние и т. д.), а не климатические или другие факторы.
Ряд специалистов объясняют колебания численности популяции тем, что в условиях перенаселенности возникает стресс, который влияет на репродуктивный потенциал, устойчивость к заболеваниям и другим воздействиям.
Влияние стресса (напряжения) рассмотрено во многих работах и, в частности, Г. Селье (1936). В качестве стресса он понимал неспецифическую реакцию (синдром) живого организма на любое сильное воздействие. Это всегда необычная нагрузка, которая не обязательно должна быть опасной для жизни, но непременно должна вызывать в живом организме "реакцию тревоги". Г. Селье выделил особые факторы - стрессоры, вызывающие ответную реакцию со стороны организма и приводящие его в напряженное, неуравновешенное состояние. Эти факторы имеют самое различное происхождение: и физическое, и биолого-физиологическое, например шум, вибрация, резкое изменение температуры или давления, радиоактивное излучение, токсины, голодание, запах и др.
При любом воздействии стрессоров в особи возникают однотипные биохимические изменения, направленные на преодоление действия этих факторов путем адаптации. Это позволяет формировать адаптационные изменения и на уровне популяции. В качестве примера рассмотрим реакцию растений на температурные воздействия. Возможно возникновение высокотемпературного стресса; при этом засухоустойчивые растения способны переносить длительное обезвоживание и перегрев, причем основным аппаратом охлаждения у растений остается транспирация. Многие суккуленты обладают способностью накапливать ночью органические кислоты (главным образом, яблочную), а днем с целью уменьшения испарения устьица закрываются, и эта кислота становится главным источником углерода, из которого синтезируются различные сахара (Жунчиету и др., 1991). Несколько иначе выглядит ситуация с возникновением низкотемпературного стресса. При отрицательных температурах образование льда в тканях растений ведет к их гибели, причем наиболее устойчивы к морозу менее обводненные организмы. Установлено, что в клеточном соке холодостойких растений северных районов содержатся растворимые сахара (глюкоза, сахароза, фруктоза), которые увеличивают концентрацию клеточного сока и уменьшают его способность к образованию внутриклеточного льда. Так, по данным В.А. Вронского (1996), количество сахара у сортов яблони в Карелии составляет 13-15%, малины - 6,5-8 % (против 5 % в средней полосе), в черной смородине - до 11 % и т. д. Это показывает, что наличие соединений в виде Сахаров способствует адаптациям растений к неблагоприятным условиям
217
Севера и таким образом пережить неблагоприятные периоды со стрессовыми воздействиями.
Комплекс стрессовых воздействий иногда вызывает стремительное падение плотности популяции, что называют "адаптационным синдромом", который препятствует излишне сильным флуктуациям, что, в свою очередь, могло бы нарушить функционирование популяции и экосистемы в целом и угрожать выживанию вида.
Наряду с вышеописанным у ряда экологов есть мнение, что изменения численности популяций определяются истощением ресурсов и уменьшением количества пищи и (или) ее питательной ценности.
Проведенное изучение динамики численности и плотности популяций выявило так называемые "всплески" плотности с уменьшающейся во времени амплитудой.
Последующее рассмотрение этих данных применительно к человеческому сообществу привело к выводу, что они должны выполняться и в том случае, если регуляция численности всей популяции или ее отдельных групп осуществляется только вследствие "демоперенаселения" (т. е. если отсутствует "внешнее" регулирование, например, планирование семьи, повышение уровня образования женщин и молодежи и т. п.). Как отмечает А.А. Горелов (1998), это особенно опасно, так как для человеческой популяции, единственной среди других известных, установлена положительная корреляция между плотностью (численностью) популяции и скоростью роста. Это особенно опасно, так как общее население людей на Земле растет, и уже превысило 6 млрд-ный рубеж, а человек по своему обыкновению мало задумывается о будущем, а действует в данный момент по данной ситуации.
Продолжая это рассмотрение, следует остановиться и на такой известной зависимости: флуктуации плотности (численности) популяции ярче выражены в сравнительно простых экосистемах, где число взаимодействующих популяций и их разнообразие также относительно мало. При уменьшении под различными внешними воздействиями видового разнообразия, а тем самым числа популяций в экосистеме возникает опасность проявления все более интенсивных флуктуации в составляющих популяциях, вплоть до их исчезновения, и этот процесс, как полагают отдельные экологи, может превратиться в необратимый. В связи с этим Ю. Одум высказывает следующее мнение: "Чем выше уровень организации и зрелости сообщества и чем стабильнее условия, тем меньше амплитуда флуктуации плотности со временем" (Ю. Одум, 1975, с. 244).
Кривые роста популяций показывают, что он может внезапно прекратиться, когда популяция близка к исчерпанию собственных ресурсов (пища, жизненное пространство), когда резко меняются климатические, эдафогенные условия и т. д. Затем при достижении внешнего предела плотность популяции может либо некоторое время оставаться на данном уровне, либо сразу же резко падать. Причем по
218
мере увеличения плотности популяции значительно усиливается действие неблагоприятных факторов среды, т. е. так называемое сопротивление среды. Рассмотренное явление получило название триггерного эффекта. Аналогичный результат был получен для характеристик, описывающих развитие мирового сообщества в результате исследований Д. Медоуза (1991).
Популяции в целом имеют тенденцию эволюционизировать таким образом, чтобы прийти к способности или уровню саморегуляции. При этом естественный отбор действует в направлении максимального повышения качества среды обитания особи и уменьшает вероятность гибели популяции.
Динамика экосистем. Функциональная целостность экосистем является их отличительной чертой и определяется сложными и межвидовыми взаимоотношениями. Эти взаимоотношения обладают некоторыми относительными "свободами" в структурных связях между отдельными компонентами, в частности это выражается в возможности замены видов в конкретных биоценозах сходными по жизненным функциям с другими видами. В зависимости от динамики численности и биологической активности отдельных популяций возможны изменения направленности и интенсивности потоков вещества и энергии, информации. Выше мы уже отмечали, что нестабильность абиотических факторов экосистемы существенным образом влияет, а зачастую и определяет колебательный характер состава и функциональных связей внутри сообществ организмов. Из этого следует, что биоценозы, являющиеся открытыми системами, непрерывно изменяются под действием внутренних и внешних факторов. Функционирование биоценозов носит вероятностный характер.
Фундаментальное свойство экосистем - это ее динамичность, которая отражает ее зависимость от комплекса факторов и представляет собой адаптивную реакцию всей системы на их воздействие.
Динамические характеристики экосистемы имеют различную продолжительность: изменения в биоценозах могут иметь суточные и сезонные колебания (ритмы); могут длиться на протяжении ряда лет и даже в пределах времени геологических эпох, выражая тем самым эволюционные процессы в биосфере, как глобальной экосистемы.
Суточные и сезонные ритмы в экосистемах. Известно, что облик биоценозов весьма изменчив и отражает комплекс адаптации экосистемы в целом к суточной и сезонной динамике условий существования. Биоценозы имеют в этом аспекте изменчивость облика, связанную с динамикой активности, с сезонными ритмами в жизни растений и животных, сменами видового состава на протяжении годового цикла. Характерной чертой такого типа динамики является сохранение принципиальных свойств данной экосистемы, поддержание ее целостности и функциональной устойчивости. Даже сезонные изменения видового
219
состава входят в общую характеристику каждой данной экосистемы, поскольку закономерно повторяются из года в год.
Суточные изменения биоценозов. В пределах суток не происходит принципиальных изменений видового состава и основных форм взаимоотношений в биоценозе, если эти изменения носят ритмичный закономерный характер. По И.А. Шилову, в данном случае имеет смысл говорить не о суточной динамике, а о суточных аспектах биоценоза. В данном промежутке времени изменения определяются характером активности тех видов, которые отличаются отчетливой суточной ритмикой жизнедеятельности. К примеру, среди рыб имеются формы с дневной и ночной активностью; известны суточные вертикальные миграции планктона и вслед за ними планктоноядных животных; достаточно широко распространены птицы с дневной и ярко выраженной ночной активностью, кбТорыё следуют за насекомыми с аналогичной суточной ритмикой, и т. п. Выявлены и некоторые экзогенные влияния на суточные ритмы, к примеру, днем в жарких пустынях снижается активность (или полностью замирает) даже тех видов, которые принципиально относятся к дневным, а некоторые из них даже меняют свой вид активности на сумеречный или даже ночной.
Суточные аспекты биоценозов отражают особенности существования в экологических нишах, в целом проявляют структурированность биоценозов по "пищевому" признаку. В частности, разделение видов, присущих одному биоценозу, по "временной" активности значительно снижает уровень прямой конкуренции и позволяет сосуществовать видам со сходными биологическими "запросами". В целом расхождение в суточной активности приводит к усложнению биоценозов, повышению биологического разнообразия и более полному использованию ресурсов среды.
Сезонные изменения (аспекты) биоценозов. Эти изменения затрагивают биоценозы гораздо глубже, чем суточные. В первую очередь это касается видового состава биоценозов. Известно, что при поступлении неблагоприятных сезонов года часть видов, для которых это имеет значение, мигрирует в районы, где условия более удовлетворяют их требованиям (перелетные птицы, некоторые копытные животные). Здесь весьма четко проявляются особенности видов, входящих в экосистему: оседлые виды (это, как правило, эдификаторы или доминанты) составляют основное ядро биоценоза, тогда как сезонные диктуют его облик. Следует сказать, что при миграционных процессах происходит изменение и характера биоценотических связей: в разные сезоны при наличии или отсутствии отдельных видов внутренние связи меняются. К числу сезонных аспектов относят и виды с пульсирующей активностью (диапаузы насекомых, оцепенение пойкилотермных и спячка гомойтермных животных и т. д.). Растительные сообщества также меняются по сезонам, как структурно (листопад, засыхание трав),
220
так и функционально (изменение фотосинтеза, накопление биомассы и т. п.).
По И.А. Шилову (2000), уменьшение числа активных видов снижает интенсивность биогенного круговорота вещества и несколько замедляет энергетический обмен, т. е. число видов в биоценозе в определенной степени следит за регуляцией биосферных процессов. Биологически значимым для сезонных аспектов является влияние абиотических факторов, особенно ярко проявляющихся в ландшафтно-климатических зонах со сменой зимних и летних периодов, а в тропиках - засушливых и влажных менее выражено.
В водной среде сезонные аспекты проявляются в связи с гидрологическими режимными сезонами, которые не совпадают в ландшафтно-климатических зонах. В качестве примера можно привести изменения видового состава планктона: "период цветения моря - биологическая весна" - массовое развитие фитопланктона, "биологическое лето" - активное развитие зоопланктона, "биологическая осень - осеннее цветение" - в умеренных климатических зонах - вспышка цветения фитопланктона, а в арктических зонах - общее снижение биомассы планктона, "зима" - время минимума биомассы планктона во всех климатических зонах.
Указанные суточные и сезонные аспекты биоценозов затрагивают, как следует из вышеизложенного, число видов, т. е. количественные характеристики, но качественного изменения системы в данном случае не возникает.
Экологические сукцессии. Динамика биоценозов может быть принципиально иной, когда под влиянием комплекса факторов меняются фундаментальные свойства экосистем и происходит их изменение в виде серии сменяющих друг друга сообществ. "Развитие биоценозов, при котором имеет место замещение во времени одного сообщества другим, называют экологической сукцессией" (от лат. sucsesio - преемственность) (И.А. Шилов, 2000, с. 432). По времени осуществления сукцессии, как правило, длятся годами и десятилетиями, хотя во временных водоемах сукцессии осуществляются с удивительной скоростью. Установлены и вековые изменения экосистем, которые отражают эволюцию биосферы.
Наиболее глубоко проблема сукцессии проработана на фитоценозах, в первую очередь потому, что смены сообществ базируются на функциях автотрофов, гетеротрофные же изменения являются вторичными и следуют за автотрофами.
Впервые динамику экосистем описал Э. Варминг в 1896 г., но наибольший вклад внес Ф. Клементе (1904). Надо сказать, что влияние Ф. Клементса на этот раздел экологии оказалось сильным, но некоторые современные специалисты, отдавая дань полученным им результатам, считают, что современные исследования уже требуют
221
переосмысления таких постулатов Ф. Клементса, как, к примеру, его утверждение о климаксовом состоянии сообщества как конечном этапе сукцессионного процесса или решающей роли климата в развитии сукцессии и ряд других.
Итак, сукцессия - это закономерная, последовательная смена сообществ в экосистемах, обусловленная влиянием комплекса внутренних и внешних факторов. Изменения во времени - это естественное свойство экологических сообществ. Влияние комплекса факторов вызываетэ экосистемах сукцессию как адаптивную реакцию. Ф. Клементc считал, что сукцессия завершается формированием сообщества, наиболее адаптированного по отношению к комплексу климатических условий, которое он назвал "климакс - формация" или просто "климакс"; в настоящее время эта формация считается временным состоянием: в процессе вековых изменений условий (климата и других факторов среды) возникают полномасштабные изменения экосистем. Выделяются прогрессивные сукцессии, в которых постепенно наращивается видовое разнообразие, но имеется также и дигрессии - регрессивные сукцессии, направленные на объединение и упрощение сообществ. Особенно часто последнее стало проявляться при наличии крупномасштабных адаптированных воздействий на биоценозы, нарушающих оптимум условий.
По современным представлениям смены сообществ могут происходить под влиянием климатических факторов, трансформации рельефа, обогащения или обеднения почв, изменения гидрологического режима и т. п. Важнейшее значение же придается биоценотическим факторам: виды растений (а также животных), участвующие в сукцессионных сообществах, изменяют условия обитания для других видов, таким образом "подготовляя почвы" для последующего этапа сукцессии. В целом выделяют сукцессии, связанные исключительно с внешними абиотическими факторами (аллогенные), и сукцессии, вызванные изменением структуры и системы связей в существующих сообществах (автогенные). Но эти два типа сукцессии способны переходить друг в друга в силу глубокой взаимозависимости (В.Н. Сукачев, 1942).
Начальный этап сукцессии выражается в процессе первоначального формирования растительного покрова. Он связан с вселением (миграцией) растений на данную территорию, их отбором в процессе приспособления к ее условиям, затем к конкуренции между ними из-за средств жизни. Это в целом приводит к формированию фитоценоза, после чего происходят структурные изменения в экосистеме, которые и приводят к устойчивому сообществу, отвечающему понятию климакс - формации, с возможными дальнейшими уже вековыми изменениями (Шилов И.А., 2000).
По Ф. Клементсу, в наиболее общем виде сукцессии проходят через фазы обнажения (появление незаселенного пространства), миграции
222
(заселение его первыми, пионерными формами жизни), эцезиса (колонизация и приспособление к конкретным условиям среды), соревнования (конкуренция с вытеснением ряда первичных поселенцев), реакции (обратное воздействие сообщества на биотоп и условия существования) и стабилизации (формирование климаксового биоценоза). В сукцессионных процессах, по мнению В.Н. Сукачева, важнейшую роль играют конкурентные отношения на внутривидовом и главным образом на межвидовом уровне, которые в конечном итоге и приводят к равновесному состоянию, характеризующему завершающее сообщество.
Сукцессия, в энергетическом смысле, связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, направленной на поддержание системы. Стадии роста, стабилизации и климакса сукцессии можно различать на основе критерия продуктивности: на первой стадии продукция растет до максимума, на второй остается постоянной, а на третьей по мере деградации системы уменьшается до нуля.
Наиболее явно различие между растущими и зрелыми системами следует из табл. 16, в которой видна обратная зависимость между энтропией и информацией, а также то, что развитие экосистем идет в направлении повышения их устойчивости.
По общему характеру сукцессии подразделяются на первичные и вторичные. Первичные сукцессии начинаются на субстрате, неизмененном (или почти неизмененном) деятельностью живых организмов. Главной функцией такого рода сукцессией является создание (или изменение) почвы первичными колонистами.
Таблица 16. Различия в экосистемах разного возраста
Показатели
Растущая экосистема
Зрелая экосистема
- Урожай
- Видовое разнообразие
- Структурное разнообразие
- Специализация по нишам
- Размеры организма
- Жизненные циклы
- Скорость обмена биогенных
веществ между организмом и средой
- Давление отбора
- Внутренний симбиоз
- Сохранение биогенных веществ
- Стадийность
- Энтропия
- Информация
- высокий
- мало
- слабо организовано
- широкая
- небольшие
- короткие и простые
- высокая
- на быстрый рост
- не развит
- с потерями
- низкая
- высокая
- мало
- низкий
- велико
- хорошо организовано
- узкая
- крупные
- длинные и сложные
- низкая
- на регуляцию
обратной связи
- развит
- полное
- высокая
- низкая
- много
223
В качестве примера остановимся на формировании скальных биоценозов: процесс начинается с поселения на скалах накипных лишайников. На этой стадии постепенно формируется комплекс видов микроскопических водорослей, простейших, нематод, отдельных насекомых, который способствует формированию первичной почвы. Затем здесь поселяются другие формы лишайников, специализированных мхов; еще позднее на базе возникшей почвы вселяются сосудистые растения и параллельно идет обогащение животного населения. Сходным образом описывается смена сообществ на ледниковых отложениях на очень бедных почвах, завершающее сообщество здесь создается примерно через 100 лет после начала сукцессии (Шилов И.А., 2000). Вторичные сукцессии развиваются на субстрате, первоначально измененном деятельностью комплекса живых организмов. Такие сукцессии чаще всего имеют восстановительный (демутационный) характер.
Примером вторичных демутационных сукцессии может служить восстановление климаксового лесного биоценоза после пожаров (а в наше время и вырубок). В таежной зоне Евразии при появлении открытого пространства на месте еловых лесов в результате пожара или сплошной рубки коренным образом меняется режим освещения, температуры, влажности и других факторов. Эти изменения неблагоприятны для таежных видов растений и животных, развитие которых угнетается. Зато на освещенных, относительно сухих и хорошо прогреваемых местах формируется временное одноярусное сообщество из светолюбивых трав. Первыми на стадии открытой вырубки развиваются виды с легко распространяемыми семенами: вейник, Иван-чай и др. Позднее начинается лесовозобновление: прорастают светолюбивые лиственные породы (осины, березы, ивы и др.) и кустарники. Одновременно формируется связанный с лугово-кустарниковой растительностью комплекс животного населения: многочисленные и разнообразные насекомые и другие беспозвоночные, грызуны, исходно связанные с лугами и лесными полянами; развитие ягодных кустарников привлекает большое число видов птиц, а наличие хорошо прогреваемых мест - рептилий и т. д. Формируется богатое и разнообразное сообщество зарастающей вырубки.
Эта стадия занимает в среднем 2-3 года, после чего начинается интенсивное развитие светолюбивых мелколиственных древесных пород (осины, березы). Постепенно подросшие деревья вытесняют кустарники и наиболее светолюбивые виды трав; кустарниково-луговое сообщество сменяется лиственным жердняком - молодым лесом с несомкнутыми кронами. Это влечет за собой и изменение животного населения: на стадии жердняков они обедняются за счет миграции видов, ранее связанных с кустарниками и богатым разнотравьем. После смыкания крон и перехода сообщества от стадии жердняка в фазу лиственного леса (через 10-15 лет от начала сукцессии) биоценоз вновь несколько усложняется за счет появления более или менее
224

Рис. 66.. Первичная сукцессия
полного комплекта растительности и животного населения, характеризующего спелые лиственные леса. Под древесным пологом в условиях затенения и повышенной влажности начинается интенсивное прорастание семян ели. Постепенно хвойный молодняк окончательно заглушает луговую травянистую растительность; ее сменяют мхи и лесное разнотравье. Улучшение условий для роста ели ингибирует
225
восстановление мелколиственных лесных пород. Старые лиственные деревья затеняются выходящими в первый ярус елями; смыкание еловых крон еще больше угнетает березы и осины, которые выступают уже в качестве второстепенных членов древостоя. В конце концов лиственные деревья выпадают и восстанавливается исходный тип лесного сообщества с господством ели. Соответственно изменяется и фаунистический комплекс, прошедший в процессе сукцессии фазы сорно-луговой растительности, кустарников, лиственного жердняка и спелого леса. Весь процесс от вырубки (пожара) до формирования устойчивого таежного биоценоза занимает от 90 до 150 лет". (И.А. Шилов, 2000, с. 436-439). В табл. 17, 18 приведены некоторые данные по фауне на различных сукцессионных стадиях (рис. 66).
Подобным образом описываются сукцессии в других зональных лесных биоценозах. Однако сукцессия не всегда доходит до восстановления исходного биоценоза, она может остановиться и на промежуточных стадиях, например пастьба скота на вырубках и пожарищах может "повернуть" сукцессию в сторону формирования суходольного луга. При значительных переувлажнениях может произойти заболачивание вырубки, что препятствует восстановлению древесной растительности. Иногда процесс может остановиться и на стадии березняка и осинника.
Таблица 17. Изменение видового состава птиц на зарастающих вырубках,
Средний Урал (по Н.Н. Данилову, 1958 г., с изменениями автора)
Таблица 18. Численность насекомых на вырубках и в лесу
(по М.Н. Керзиной, 1956 г., с изменениями автора)
Место учета
Абсолютное количество
- Открытые вырубки
- Зарастающие вырубки
- Спелый лес
Примечания: 1. По исследованиям в Мордовском заповеднике (сосновый лес с березой, елью, осиной). 2. На 100 взмахов сачком со сбором насекомых (цикад, клопов, жуков, пауков).
К вторичным сукцессиям относят и такие, при которых исходной силой, вызывающей смену сообществ, оказываются нарушения
226
стабильных взаимодействий в биоценозе; это, например, зоогенные сукцессии, выражающиеся в изменении фитоценоза при чрезмерном выпасе скота; на пастбищах увеличивается число копрофагов, специфических паразитов домашнего скота, а также концентрируются крупные хищники и падальщики. Представляется интересным изложение следующего биологического факта (И.А. Шилов, 2000, с. 441-442): "В середине нашего столетия в лесной зоне Восточной Европы в результате направленной охраны существенно возросла численность лосей Alces alces. Этому в немалой степени способствовали широкомасштабные рубки леса и связано с этим появление площадей, зарастающих лиственными и сосновыми молодняками. Лоси скапливались в таких местах, в результате чего нарушался естественный ход сукцессии, на больших площадях погибал подрост сосны, осины, дуба, ивы и других пород. Это отрицательно сказывалось на восстановлении лесных насаждений и вызывало глубокие изменения в состоянии и динамике лесных биоценозов".
Выделяются также деградационные сукцессии, заключающиеся в последовательном использовании различными видами разлагаемой органики. Особенность деградационных смен заключается в том, что такие сообщества составлены только гетеротрофными организмами, а направление сукцессии идет в сторону все большего структурного и химического упрощения скоплений органического вещества. Это выражается в изменении видового состава насекомых и других организмов, потребляющих древесину на разных стадиях ее разложения. Аналогичная динамика имеет место в скоплениях помета животных (серии смен копрофагов), в разлагающихся растительных остатках, а также в сильно загрязненных органикой водоемах. Образующиеся в этих условиях временные сообщества не соответствуют понятию полного по структуре и составу биоценоза и виды в них объединяет только общность пищевого субстракта, а серии сменяющих друг друга наборов видов функционально относят к цепям редукции.
Вековые смены экосцстем. Фундаментальные крупномасштабные формы динамики биоценозов представлены вековыми сериями сменяющих друг друга экосистем на протяжении многих столетий. Эти экосистемы представляют собой устойчивые ландшафтные сообщества. Масштабность изменений в экосистемах такого рода в целом сопровождает и определяет развитие биосферы в целом. Эти сукцессии охватывали по длительности целые геологические периоды и осуществлялись как смены типов сообществ живых организмов в связи с изменениями климата, рельефа и других свойств геосфер Земли.
В целом такие сукцессии не являются типично экологическими в их классическом выражении. Дело в том, что смены флоры и фауны, протекающие в геологических масштабах, начинаются отнюдь не с заселения "пустырей" (не занятых жизнью пространств), а с перестройки внутренних связей уже сложившихся и функционирующих экосистем.
227
По И.А. Шилову (2000), А.А. Горелову (1998), причиной смены видового состава в этом случае оказывается то обстоятельство, что медленно идущие изменения среды приводят к тому, что ряд видов теряет ранее выработанную приспособленность к условиям обитания; они замещаются другими видами, более адаптированными к новым условиям. Самым типичным образцом такого рода сукцессии является изменение фитоценозов и зооценозов по мере отступления ледников в периоды межледниковий. Следует заметить, что трансгрессия и регрессия ледников иногда столь значительно трансформировала почвенный покров, что в ряде случаев было необходимо собственно новое почвообразование, и сукцессия превращалась в типичную для первичного типа.
К числу вековых сукцессии можно отнести процессы, протекающие в Аральском регионе (Палваниязов, 1992, Шилов, 2000). Здесь необходимо сделать существенное замечание: исходная причина изменений в экосистеме данного региона связана в наибольшей степени с хозяйственной деятельностью человека, которая нарушила сложившийся водный баланс небольшого по площади Аральского моря.
Примерно до 1960 г. Аральское море оказывало серьезное смягчающее влияние на "континентальность" климата прилегающих и довольно значительных по площади территорий. Поглощая тепло летом и отдавая его в окружающие пустыни зимой, море смягчало температурные экстремумы и стабилизировало тем самым условия жизни биоценозов Приаралья. Испаряемая морем влага уменьшала сухость воздуха и частично восполняла дефицит влаги, которая становилась доступной живым организмам. С начала 60-х годов началось прогрессирующее снижение уровня Аральского моря: к концу 70-х годов площадь его акватории сократилось на 16 тыс. км2, к 1985 г.- на 19 тыс. км2, а затем ежегодно с каждым метром снижения уровня моря обнажалось до 2 тыс. км2 его дна. Этот процесс существенно изменил гидрогеологическую обстановку на значительных по площади территориях: снижение уровня грунтовых вод в устье Аму-Дарьи составило 4 м, а выше по ее течению в Кызылкумах до 6-11 м. Обсыхающее дно моря характеризуется интенсивным процессом опустынивания, в котором ведущими факторами становятся поверхностное засоление почвы и ветровые (эоловые) процессы - дефляция и десквамация, возникает значительный перенос частиц соли. Дно моря заполнено песчано-солончаковыми массивами. Эти активные изменения в абиотических факторах среды сказываются на состоянии экосистем прилежащих к бывшему побережью территории в полосе более чем 300 км. Солончаково-песчаные равнины вначале - на второй год после обнажения - колонизируются солянками. Сюда вселяются песчанки, малый тушканчик, домовая мышь; вслед за ними появляются некоторые хищники (ласка, лисица, степной хорь). Появляются и копытные - кабан, местами - сайгак, джейран. Но уже через 3-4 года эти места
228
превращаются в так называемые пухлые солончаки, лишенные растительности и животного населения. Повышение уровня грунтовых вод влечет за собой расширение зоны сыпучих песков; в климате происходит постепенное наращивание степени "аридизации". Все это сказывается и на видовом составе численности и условиях существования живых организмов на прилежащих территориях Кызылкумов, Приаральских Каракумов, плато Устюрт и др.
Гомеостаз на уровне экосистем. Многообразные формы связей, существующие между популяциями разных видов, объединяют биоценоз в целостную биологическую систему. Как все биологические системы любого уровня, биоценоз существует в определенных абиотических условиях того или иного участка географической среды. Эта среда никогда не является абсолютно стабильной, а устойчивое выполнение главенствующей функции экосистем - поддержания биогенного круговорота - должно обеспечиваться биоценотическими адаптивными механизмами. Эти механизмы при рассмотрении их на уровне экосистем носят исторический характер, так как складываются на протяжении всего существования и формирования экосистем. Между элементами экосистемы устанавливаются общие стабильные взаимоотношения, которые адаптированы не только к общим факторам существования данной экосистемы, но и к степени взаимосвязей, определяющих устойчивое поддержание этих отношений в колеблющихся условиях данной конкретной среды.
Любая экосистема любого иерархического уровня может устойчиво функционировать только в пределах устойчивой реализации обратных связей или в области нарушения этих связей, когда элементы экосистемы способны компенсировать отклонения, определяемые положительной обратной связью (например, при поступлении загрязнений в водную экосистему она еще способна к самоочищению). Эта область устойчивости экосистемы называется гомеостатическим плато (см. рис. 65). В пределах (верхнем и нижнем) действия обратных связей экосистема за счет компенсаторных регуляторов сохраняет устойчивость. В антропогенных экосистемах при возникающих соответствующих нагрузках для устойчивого их функционирования человек должен сам играть роль компенсаторного регулятора (озеленение, посадка леса, системы очистки воздуха и воды).
Нарушения в передаче информации по каналам обратной связи возникают в результате стихийных бедствий: засух, наводнений, землетрясений, болезней. В результате человеческой деятельности также возникают помехи в реализации обратной связи - застройка городов, изменение ландшафта и т. п. Эти нарушения как природного, так и техногенного характера носят иногда случайный характер и они играют роль помех, не разрушая функционирование экосистемы.
На уровне экосистем перечень видов, состав и сложность развития трофической сети, наиболее устойчивые формы взаимодействия между
229
видовыми популяциями отражают приспособленность к наиболее определяющим особенностям среды и направлены, прежде всего, на устойчивое поддержание биогенного круговорота в изменяющихся условиях. Нарушения, которые возникают в экосистемах на фоне установившихся средних параметров среды, вызывают функциональные адаптации компенсаторного типа. При этом сохраняется принципиальная структура биоценоза. Это многообразные обратимые изменения пищевых цепей, паразитарных связей, условий среды, обусловленные колебаниями численности (плотности) популяций, выражающихся на уровне видов. При более существенном нарушении состава биоценоза возникают неустойчивые, сменяющие друг друга сообщества - процесс, в идеальном, кстати весьма редком, случае ведущий к восстановлению исходного типа экосистемы. Экологические сукцессии такого рода - одно из наиболее ярких выражений действия функциональных адаптации на уровне биоценозов.
Если изменения среды имеют необратимый характер или отмечена устойчивая тенденция к приобретению средой именно такого характера, то происходит направленная смена типов сообществ. В целом регулируется смена уровня стабилизации биоценотической системы. Особенно точно это прослеживается при оценках в масштабах геологического времени, что нами уже отмечалось при описаниях изменения таких абиотических факторов среды, как климат, параметры рельефа, трансгрессии моря и его гидрологические режимы на протяжении таких значительных отрезков истории Земли, как геологические периоды и даже эры. Влияние этих факторов на динамику типов биоценотических сообществ уже достаточно точно установлено палеонтологическими и палинологическими исследованиями и подтверждает его наличие уже в весьма отдаленные геологические эпохи. В частности, показано, что специализированная охота палеолитического населения на крупных травоядных млекопитающих (слоны, мамонты, носороги и др.) стала причиной резкого снижения их численности и при наложившихся факторах потепления климата в межледниковьях привела к почти полному их исчезновению. Это послужило причиной коренного изменения общего облика фитоценозов. В плейстоцене крупные травоядные выступали в качестве эдификаторов (основателей) биоценозов, создавая мозаичность растительного покрова путем постоянного повреждения сомкнутых древостоев и сплошных массовых кустарников и поддержания на их месте ассоциаций злаков и разнотравья. С гибелью мамонтовой и подобной фауны сомкнутые древостой стали широко распространяться, и растительный покров Земли принял близкий к современном/зональный облик (С.П. Маслов и др., 1995).
Уже исследованиями В.Н. Сукачева было показано, что у нашей планеты имеется целостный биогеоценотический покров Земли. Действительно, хотя биологический круговорот может быть завершен уже на уровне отдельной экосистемы и даже биоценоза, в реальных
230
условиях обособленных круговоротов не установлено, да они, вероятно, и неосуществимы. На уровне биосферы круговоротные процессы объединяются в единую систему глобальной функции живого вещества. По И.А. Шилову (2000), в этой системе не только полностью завершаются отдельные биогеохимические циклы, но и реализуются тесная взаимосвязь их с абиотическими процессами формирования и переформирования горных пород, становления и поддержания специфических свойств гидросферы и атмосферы, почвообразования, поддержания их естественного плодородия и т. д. В конечном счете многообразие форм жизни определяет уникальные свойства биосферы как самоподдерживающейся системы, гомеостаз которой осуществляется (т. е. запрограммирован) на всех уровнях организации живой материи. Теснейшая функциональная взаимосвязь биологических систем разных уровней превращает отдельные формы жизни в интегрированную глобальную систему. Различные уровни гомеостазирования биологических систем и биосферы в целом сложились на протяжении длительной геологической истории нашей планеты.
231

