Глава 10
Уровни организации живого вещества
Продуценты, консументы, редуценты во всем своем многообразии, действуя совместно, определяют устойчивое поддержание глобального биологического круговорота веществ и управляют потоками энергии и информации в биосфере нашей планеты. Все составляющие этого процесса связаны пространственно-функциональными взаимоотношениями, которые выступают как стабилизаторы ее состояния при значительной изменчивости внешних и внутренних факторов. Поэтому биосферу необходимо рассматривать как глобальную экологическую систему, обеспечивающую устойчивое поддержание жизни в ее планетарном проявлении.
Экосистема (экологическая система) - это совокупность совместно обитающих разных видов организмов и условий их существования, находящихся в закономерной взаимосвязи друг с другом. Термин был введен английским экологом А. Тенсли в 1935 г. Термин "экосистема" при рассмотрении его с позиций понятий о системах представляется как "множество элементов, находящихся в отношениях и связях друг с другом, образующих определенную целостность, единство" (СЭС, 1980, с. 1225), и полностью этим понятиям отвечает. Биосфера как совокупность конкретных экосистем в ее структуре и функциях также отвечает биологическим (экологическим) системам, характеризующимся специфической функцией, упорядоченными взаимоотношениями составляющих ее частей (субсистем) и основывающимися на этих взаимоотношениях регуляторными механизмами, определяющими целостность и устойчивость системы на фоне колеблющихся внешних условий.
Выделяют микроэкосистемы (например, лиственный опад одного дерева и др.), мезоэкосистемы (пруд, небольшая роща и др.), макроэкосистемы (континент, океан) и, наконец, глобальная экосистема - биосфера Земли, которая нами уже достаточно подробно рассмотрена выше (рис. 37).
Биогеоценоз. В 1944 г. В.Н. Сукачевым был предложен термин "биогеоценоз", который не является полным синонимом экосистемы. Так, в ряде работ под биогеоценозом понимают сообщество растений, животных, микроорганизмов на определенном участке земной поверхности с его микроклиматом, геологическим строением, ландшафтом, почвой, водным режимом. Таким образом, экосистема-понятие более широкое, так как биогеоценоз - это только наземное образование с определенными границами (рис. 38).
В связи с тем, что далее мы будем употреблять ряд типичных экологических терминов, есть необходимость дать определения некоторых из них:
151
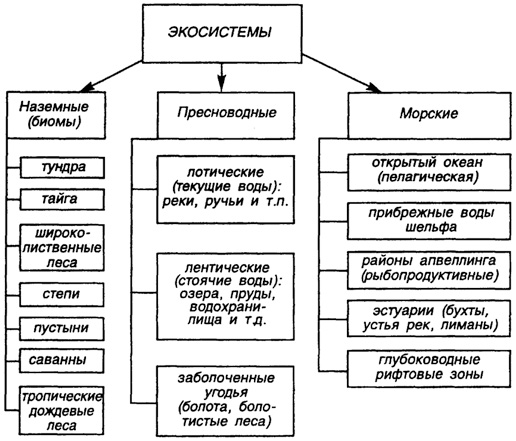
Рис. 37. Основные типы природных экосистем
- биоценоз - сообщество совместно живущих организмов;
- биотоп - среда (место) обитания;
- фитоценоз - сообщество растений;
- зооценоз - сообщество животных;
- микробиоценоз - сообщество микроорганизмов;
- биота - совокупность всех живых организмов. Соотношение между экосистемами и биогеоценозом может быть
представлено следующим образом: биогеоценоз - это экосистема в границах фотосинтеза, т. е. эти категории совпадают лишь на уровне растительного сообщества и принципиально расходятся как выше, так и ниже этого уровня. В.И. Вернадский для этих категорий использовал понятие "биокосное тело".
Ю. Одум (1986) выделяет три группы природных экосистем: наземные (биомы), пресноводные и морские. В основе подразделения лежат некоторые общие для них признаки: для наземных - тип растительности, для пресноводных - физические свойства воды и т. п.
Выделение в ландшафте различных экосистем производится достаточно произвольно, так как природные четкие границы между ними встречаются исключительно редко. Обычно между экосистемами и биомами находится переходная зона с видами и особенностями, свойственными обеим соседствующим системам. Отмечено также, что при
152
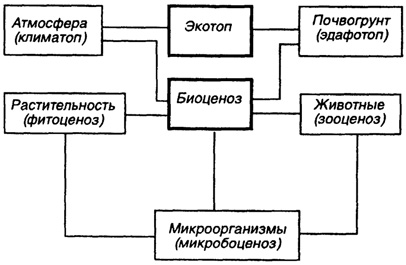
Рис. ЗЗ.Схема биогеоценоза (по В.Н. Сукачеву)
таком контакте обусловливается возникновение специфических условий среды, что в свою очередь позволяет существовать здесь особым видам растений и животных, которые присущи только этой переходной зоне. В этом случае данная зона может рассматриваться как отдельная экосистема. Ни одна экосистема не существует изолированно, а, как было показано выше, только при условии единства и целостности биосферы.
Биогеоценозы характеризуются стратиграфическим разделением на несколько ярусов разной высоты: растительность подразделяется по высоте на мхи, травянистые растения, подлесок, деревья; животные, например птицы,- на гнездящихся на поверхности земли, в стволах (дуплах) деревьев, в кронах кустарника и деревьев. В каждом стратосе (ярусе) все живые организмы взаимосвязаны, и эта связь горизонтального типа внутри биогеоценозов носит название синузии (рис. 39). Рассмотрение видового состава биогеоценозов позволило выявить среди населяющих их живых организмов следующие:
- эдификаторы - виды, которые играют роль основоположников сообщества, т. е. живые организмы, определяющие особенности биоты данного биогеоценоза в его конкретных абиотических условиях; виды, начавшие освоение данной территории и создавшие условия для формирования трофической сети;
- доминанты - виды, которые преобладают в данном биогеоценозе, имеют наибольшую численность и плотность популяции;
- предоминанты - виды, которые живут за счет доминантов, включая внутривидовые и межвидовые отношения (паразитизм, симбиоз, мутуализм) и отношения по трофическим цепям.
Весьма важным фактором, определяющим особенности функционирования растительных сообществ, является их водообмен с окружающей средой. По виду путей регуляции водообмена выделяют:
- гидратофиты - водные растения, целиком или большей своей частью погруженные в воды (кувшинки, водяные лютики, рдест, уруть и др.);
- гидрофиты - наземно-водные растения, погруженные в воду только своей нижней частью и, как правило, растущие по берегам водоемов, на болотах или на мелководьях (тростник обыкновенный, частуха, калужница болотная и др.);
153
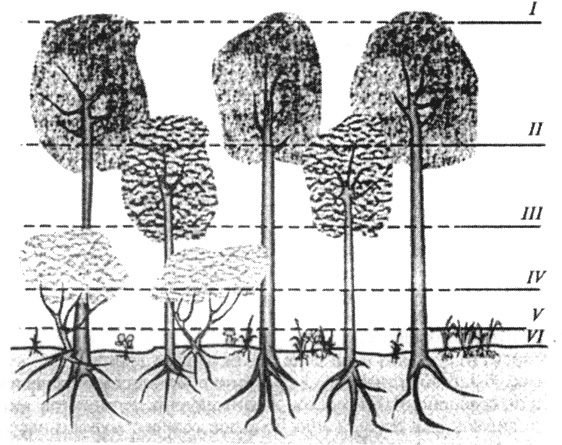
Рис. 39. Ярусность в биоценозе
- гигрофиты - наземные растения, распространенные в зонах повышенной влажности и, как обязательные условия, на влажных почвах; среди них различают теневые гигрофиты:- это растения нижних ярусов сырых лесов в разных климатических зонах (недотрога, бодяк огородный, подавляющее большинство тропических трав и др.); световые гигрофиты - встречающиеся, как правило, на открытых местообитаниях (рис, папирус, росянка, подмаренник болотный и др.);
- мезофиты - растения, которые могут переносить не очень сильную и непродолжительную засуху, но произрастание их обычно приурочено к среднему увлажнению и умеренно теплому климату; они занимают промежуточное положение между гигрофитами и ксерофитами; в эту группу входят травянистые растения умеренной зоны (пырей ползучий, клевер луговой, люцерна синяя), большинство культурных растений (кукуруза, пшеница, горох, конопля, соя и т. п.); почти все плодовые (за исключением винограда, миндаля и фисташки), а также листопадные деревья саванн, кустарники подлеска и др.;
- ксерофиты - это растения, приспособленные к жизни в местообитаниях с недостаточным увлажнением; растения в лучшей степени, чем другие, способны регулировать водообмен, имеют приспособления, уменьшающие транспирацию (узкие листья, толстая кутикула, опущение, редукция листьев и др.); к ним относят, главным образом, растения степей, полупустынь, пустынь, песчаных дюн и т. п. (представители семейств - маревых, сложноцветных, крестоцветных, бобовых, гвоздичных, лилейных, парнолистников и др.); они, в свою очередь, подразделяются на два основных типа:
154
- а) суккуленты - сочные растения, обитающие в засушливых районах и переносящие неблагоприятный для жизнедеятельности период за счет накапливаемых запасов влаги в своих тканях,- это могут быть стебли (кактусы, кактусовые молочаи), листья (агавы, алоэ); при этом кактуеовидная форма растений, принадлежащих другим семействам (молочайных, сложноцветных), также служит приспособлением к аналогичным засушливым условиям внешней среды; из деревьев, которые запасают воду в стволах, более всего известны африканский баобаб, австралийское бутылочное дерево, произрастающие в саваннах; по некоторым данным (Ф. Фукарек и др., 1982) африканский баобаб при общей высоте дерева до 20 м способен запасти в своей мягкой древесине до 120 тыс. литров влаги;
- б) склерофиты - это растения, в отличие от суккулентов, имеют "сухой" внешний вид с жесткими, кожистыми листьями и стеблями, которые эффективно задерживают испарение воды (полынь, саксаул, олеандр, некоторые злаки, оливковое дерево, синеголовник полевой и др.), распространены преимущественно в засушливых районах (рис. 40).
Водные экосистемы имеют свои особенности и также стратифицированы, например разные животные, растения и даже микроорганизмы обитают на различной глубине. Строение водных экосистем определяется характером водной среды: непроточная (лентическая); проточная (лотическая). В целом же водные организмы в этих экосистемах в зависимости от мест обитания подразделяют на:
- бентос (от греч. bentos - глубина) - донные организмы, обитающие на дне водоема; в независимости от его глубины могут быть прикрепленными, сидячими (кораллы, водоросли, губки, мшанки), роющими (кольчатые черви, моллюски), ползающими (ракообразные, иглокожие) или функционально связанными с условиями жизни у дна водоема (брюхоногие моллюски, скаты и др.). Наиболее богаты бентосом прибрежные районы океанов и морей, особенно северо-западная часть Атлантического и Тихого океанов;
- перифитон (от греч. peri - около, fitonos -растение) - организмы, поднимающиеся от дна к поверхности водоема, либо прикрепленные к вышеуказанным, например к стеблям растений;
155
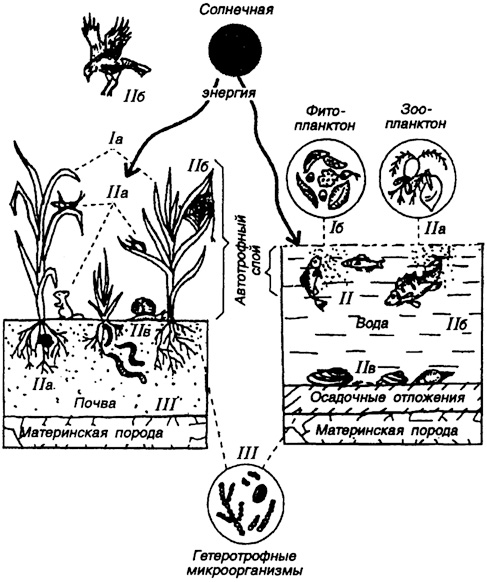
Рис. 40. Сравнение общей структуры наземного и водного биоценозов растения: Ia - высшие; Iб - водоросли; животные: IIа - растительноядные; IIб - плотоядные, IIв - всеядные; III - микроорганизмы
- планктон (от греч. planktos - парящий) - совокупность пассивно плавающих и переносимых морскими течениями в толще воды бактерий (бактериопланктон), животных (зоопланктон), растений (фитопланктон), которые почти или полностью неспособны к самостоятельному передвижению на сколько-нибудь значимые расстояния. Одной из важнейших задач жизнедеятельности этих организмов является поддержание плавучести. Этого они добиваются двумя способами: уменьшением массы тела и увеличением силы трения тела по отношению к водной массе. Это является одним из видов морфологической адаптации к абиотическим фактором среды. Для снижения массы тела, замедляющей погружение, наилучшим решением является собственно уменьшение размеров организмов, поэтому фитопланктон, в частности, представлен микроскопическими одноклеточными водорослями: диатомовыми и перединиевыми (дианофлягелляты). Другой способ, который повышает силу трения, выражается в увеличении площади поверхности тела, например, за счет уплощения формы (дисковидный диатомеи) или при помощи выростов, отростков (шипы у ракообразных, иглы у радиолярий и др.). Зоопланктон в этом отношении имеет еще больше разновидностей форм тела организмов (одноклеточные, рачки, черви, медузы и др.).
156
Фитопланктон - основной первичный продукт органического синтеза в морских водоемах, за счет которого существуют гетеротрофные водные организмы. С фитопланктоном связаны такие явления в океане, как "цветение вод" и "красные приливы";
- криль - это мелкие морские ракообразные (бокоплавы, креветки - эвфузиты и др.), служащие основным источником пищи китообразных. В частности, подсчитано, что взрослый кит за летний сезон (продолжительностью около 150 дней) потребляет около 120 т криля. До недавнего времени выделение этих разновидностей организмов в водных экосистемах носило сугубо промысловое значение, но сейчас его применяют и для различного рода экологических исследований;
- нектон (от греч. nektos - плывущий) - это группа свободно плавающих организмов, которые могут преодолевать силу течения воды, перемещаться на значительные расстояния и преодолевать силу гравитации, поднимаясь или опускаясь в толще воды от дна до поверхности; к числу этих организмов относят различных рыб, китообразных, тюленей, морских змей, черепах, кальмаров, осьминогов и др. Так, в Мировом океане насчитывается около 16 000 видов рыб; численность антарктических китов (финвал, синий кит, горбач, кашалот и т. п.) составляет не многим более 750 тыс. особей, а дельфинов - примерно 425 млн. особей и т. д.;
- плейстон (от греч. pleistos - плавание) - совокупность гидробионтов, держащихся на поверхности воды или полупогруженных (физалия или португальский кораблик, сифонофоры и др.);
- нейстон (от греч. neistos - плавающий) - сообщества организмов, обитающих в зоне пленки поверхностного натяжения воды (над или под ней); в свою очередь, подразделяется на гипонейстон - живут непосредственно под пленкой (личинки кефали, хамсы, веслоногие рачки, саргассовый кораблик) и эпинейстон - обитают над пленкой (клопы-водомерки, жуки-вертячки, из растений - сальвиния плавающая и др.).
Все рассмотренные экологические группы морских организмов участвуют в круговороте "жизни" в океане (рис. 41). Солнечная энергия, поглощаемая растениями, передается от них животным и микроорганизмам в виде потенциальной энергии по основным трофическим цепям. Эти группы консументов обмениваются с растениями диоксидом углерода и минеральными питательными веществами.
Одни и те же виды растений и животных, а тем более микроорганизмов, встречаются в весьма различных и удаленных друг от друга экосистемах.
Основной функцией экосистем (биогеоценозов) является поддержание в устойчивом состоянии биогенного круговорота. Эта функция основана на пищевых взаимодействиях, выстроенных в четко упорядоченные зависимости.
Биогеоценоз включает в себя как совокупность на определенной
157
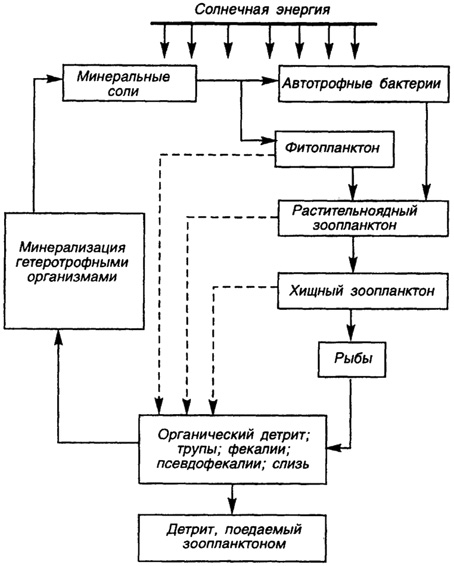
Рис. 41. Упрощенная схема круговорота жизни в Мировом океане
(по Ж. Эрхарду и Ж. Сежену, 1984)
территории, кроме биоценоза (биотических факторов - зооценоза, микробиоценоза, фитоценоза), экотоп (абиотические факторы). К числу абиотических факторов относят:
- климатические - свет, температура, влага, движение воздуха, атмосферное давление;
- эдафогенные (от греч. "edafos - почва) - механический состав влагоемкость, воздухопроницаемость, плотность альбедо;
- орографические - рельеф, высота над уровнем моря, экспозиция склона;
- химические - газовый состав воздуха, солевой состав воды, концентрация, кислотность и состав почвенных растворов.
Перечисление факторов неживой природы, оказывающих непосредственное влияние на функционирование биогеоценоза, можно продолжить, но даже того, что указано, достаточно для осознания сложности происходящих процессов.
Как пишет А.Н. Тетиор (1992), все компоненты биогеоценоза
158
взаимозависимы и взаимосвязаны. Так, климат влияет на состояние почвы и подстилающего грунта, а от состояния почвы зависит в определенной степени ее отражательная способность (альбедо), нагревание. Живые организмы являются для других живых организмов источником пищи, средой обитания или фактором смертности; они связаны между собой. Биогеоценоз - это часть планетарной экосистемы, внутри которой происходит передача информации между отдельными компонентами, осуществляется круговорот веществ и движется поток энергии.
При изучении биотической структуры экосистем становится очевидным, что к числу важнейших взаимоотношений между организмами относятся пищевые биотические факторы - совокупность влияния жизнедеятельности одних организмов на другие, характеризуемая взаимоотношениями между животными, растениями, микроорганизмами. Эти взаимоотношения носят название коакций - прямых и косвенных.
Абиотические факторы среды - это компоненты и явления неживой, неорганической природы, прямо или косвенно воздействующие на живые организмы. Естественно, что эти факторы действуют одновременно и значит, что все живые организмы попадают под их влияние. Степень присутствия или отсутствия каждого из них существенно отражается на жизнеспособности организмов, причем на разные их виды неодинаково. Надо отметить, что это очень сильно влияет на всю экосистему в целом, на ее устойчивость.
Понятно, что при колоссальном многообразии живых организмов их виды развиваются наиболее активно в разных условиях. Для одних нужна повышенная влажность и температура, для других - сухость и высокая температура и т. д. Для каждого из нас хорошим примером может служить достаточно обычный жизненный факт, а именно комнатные растения и уход за ними. Все знают, что они развиваются лучшим образом, если количество поливов их водой носит определенный характер: как перерыв в поливах, так и излишнее количество воды приводит к угнетению комнатных растений, а иногда и к гибели. Аналогичные данные получены по освещенности и температуре для комнатных растений и для животных, растений и микроорганизмов в "дикой природе". Полученные материалы, если изобразить их графически, показывают (рис. 42), что на графике имеется точка, при которой наблюдается максимальное развитие, называемое оптимумом, в некотором же диапазоне от этой точки условия для развития организмов остаются достаточно благоприятными, поэтому вполне возможно говорить о выделении некоторой зоны оптимума. При этом весь диапазон условий от их минимальных значений до максимальных, при которых возможно развитие организмов, называют диапазоном устойчивости. Между зоной оптимума и пределами устойчивости, при приближении к которым организмы испытывают постепенное угнетение, выделяются
159
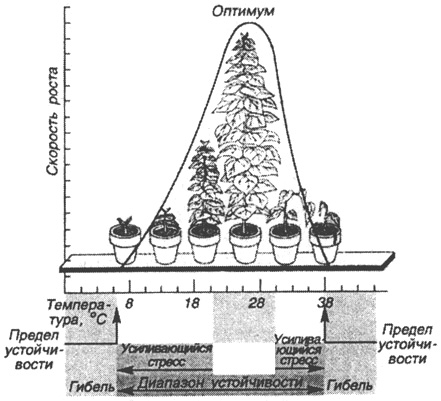
Рис. 42. Оптимум факторов воздействия на организмы
(по Б. Небелу, 1992)
зоны стресса в рамках диапазона устойчивости. Как показали многочисленные исследования, данные результаты наблюдений для подавляющего вида живых организмов указанная зависимость сохраняется при естественном различии влияния абиотических факторов на разные виды. Однако, если оптимумы и пределы устойчивости у разных видов неодинаковы, их общие диапазоны устойчивости могут в значительной степени перекрываться.
Установленные зависимости относятся к категории важнейших закономерностей в экологии. Надо сказать, что исторически первым для этой науки был закон, устанавливающий зависимость живых систем от факторов, ограничивающих их развитие (так называемых лимитирующих факторов).
Закон минимума. Ю. Либих в 1840 г. установил, что урожай зерна зачастую зависит не столько от питательных веществ, которые ему необходимы в ощутимых количествах, а сколько от тех, которых нужно совсем немного, но их явно недостаточно в почвах. Сформулированный им закон гласил: "Веществом, находящимся в минимуме, управляется урожай и определяется величина и устойчивость последнего во времени". Этот закон подтвердился, как мы выше указывали, не только избытком или недостатком питательных веществ, но и температурой, влажностью, освещенностью и т д. Действия закона ограничивают два принципа. Первый: закон Либиха строго применим только в условиях стационарного состояния. Поэтому более точной формулировкой
160
будет: "при стационарном состоянии лимитирующим будет то вещество, доступные количества которого наиболее близки к необходимому минимуму". Второй же принцип касается взаимодействия факторов среды. Высокая концентрация или доступность некоторого вещества может изменить необходимость потребления минимального питательного вещества. Многие организмы способны заменить одно дефицитное вещество другим, имеющимся в избытке (А.А. Горелов, 1998). Дальнейшие разработки позволили сформулировать так называемый закон толерантности (от лат. tolerantis - терпение): "отсутствие или невозможность развития экосистемы определяется не только недостатком, но и избытком любого из факторов (тепло, свет, вода)". Значит, организмы характеризуются как экологическим минимумом, так и максимумом. Как говорится, "слишком много хорошего тоже плохо". Диапазон этот составляет пределы толерантности, в которых организм нормально реагирует на влияние среды. Закон толерантности предложил В. Шелфорд в 1913 г. В целом же "толерантность - это способность организма переносить неблагоприятное влияние того или иного фактора среды" (СЭС, 1980, с. 1348). Сформулированы также некоторые развивающие положения этого закона:
- живые организмы имеют широкий диапазон толерантности в отношении одного фактора и в то же время узкий в отношении другого;
- широкий диапазон толерантности по всем факторам позволяет организмам, которые им обладают, распространяться более широко;
- в случае если условия по одному экологическому фактору не являются оптимальными для вида, то это влечет за собой сужение диапазона толерантности к другим экологическим факторам;
- в природных условиях организмы почти всегда оказываются под воздействием не оптимальных по значениям факторов, которые установлены для этих организмов в лабораторных условиях;
- период размножения обычно является критическим; в этот период многие факторы среды часто оказываются если не лимитирующими, то очень близкими к ним.
Мы уже отмечали, что живые организмы способны к некоторому изменению условий среды с целью ослабления лимитирующего влияния ее факторов. Виды, которые имеют широкое географическое распространение, образуют адаптированные к местам обитания свои общности, называемые экотипами. Оптимумы экотипов и пределы толерантности полностью отвечают конкретным местным условиям. В этом случае, если экотипы закреплены в генетической информации, можно говорить об образовании генетических рас или о простой физиологической акклиматизации. Кстати, акклиматизация - это приспособление живых организмов к новым условиям существования, к новым биоценозам. Акклиматизация бывает природной (миграции животных, перенос семян растений животными в новые места и т. п.)
161
и искусственной (после интродукции животных и растений). Акклиматизированные организмы могут в новых условиях давать жизнеспособное потомство (СЭС, 1980, с. 31).
Границы распространения организмов - ареал - обусловлены соблюдением необходимых требований данного организма к условиям (факторам) среды. Каждый вид организмов формирует его из требований к территории, к различным абиотическим факторам, пище, воспроизводству и другим функциям организмов. Вся эта совокупность различных параметров среды - абиотических и биотических факторов, - необходимых для оптимального обитания и развития вида, определяет экологическую нишу. Чрезвычайно важным является следующая установленная закономерность: все факторы в экологической нише взаимозависимы, взаимообусловлены, изменение одного из них влечет за собой изменение других, не подвергающихся воздействию.
Экологическая ниша представляет собой место вида в экосистеме, определяемое его биотическим потенциалом и совокупностью факторов внешней среды, к которым он приспособлен (В.А. Вернадский, 1996). Один же из самых крупных экологов современности Ю. Одум определяет ее следующим образом: "экологическая ниша - это не только физическое пространство, занимаемое организмом, но и функциональная роль организма в сообществе (его трофическое положение) и его место относительно градиентов внешних факторов - температуры, влажности, рН почвы и др." (Ю. Одум, 1986, т. 2, с. 119). Экологическая ниша как пространственное формирование характеризуется прежде всего шириной ниши и перекрыванием ниши с соседними.
Представление об экологической нише было впервые высказано зоологом Дж. Гринеллом (1914), который рассматривал ее в большей степени как пространственное образование сродни арелу, хотя и придавал значение и биологическим потребностям видов в этом пространстве.
У. Элтон (1927) под экологической нишей подразумевал в гораздо большей степени тип питания, т. е. место в трофических цепях. Практически отвечающими данному выше определению экологической нише были разработки Дж. Хатчинсона (1957), который впервые рассмотрел ее как сумму всех связей организмов данного вида с абиотическими условиями среды и с другими видами организмов. В этом случае обойтись лишь шириной ниши и ее перекрытием можно только для самых приближенных оценок, в целом же необходимо рассматривать экологическую нишу как многомерное пространство, по каждой из ординат которой располагаются пределы требований вида, и отдельным экологическим факторам. Объем экологической ниши определяется как требованиями вида к среде, так и степенью реакции среды на эти требования. В первом случае - это фундаментальная экологическая ниша, а во втором, когда среда не всегда (скорее,
162
зачастую) в полном объеме удовлетворяет требованиям, - это реализованная ниша. Перекрытие же одной ниши другой тесно увязывается с конкурентными отношениями организмов, но это тоже может быть оценено как одна из ординат "экологического гиперпространства".
Надо отметить, что, согласно Ю. Одуму (1975), экологическая ниша некоторого организма зависит не только от того, где он живет, но и от того, что он делает (как он преобразует энергию, каково его поведение, как он реагирует на физическую и биологическую среду и как он ограничен другими видами). Этот исследователь определил экологическую нишу как "профессию" вида в обществе. Специализация вида по питанию, использованию пространства, времени активности и другим условиям влияет на динамику и размеры экологических ниш. Даже близкородственные виды, живущие совместно, специализируются по питанию весьма тонко, например копытные в саваннах Африки просто разделяют между собой растение, намеченное в пищу: зебры срывают верхушки трав, антилопы гну питаются оставшимися листьями после зебр, газели выщипывают низкую траву, а антилопы топи доедают сухие стебли.
Очень близко к рассмотрению экологической ниши находится понятие жизненных форм различных видов. Жизненную форму виды приобретают в процессе эволюции при жизни в тех или иных условиях.
Жизненная форма - это комплекс видовых приспособлений, обеспечивающих успешное выживание и воспроизведение вида в данных условиях. Жизненные формы могут быть весьма схожими у разных видов, живущих в одинаковых экологических нишах, что часто выражается даже в морфологическом сходстве представителей совершенно разных неродственных видов. Классический пример такого сходства "ихтиозавр - тунец - дельфин", которое объединяет быстро плавающих в водной среде (И.А. Шилов, 2000). Изучение жизненных форм представляет собой весьма сложную, комплексную проблему в связи со значительным разнообразием и комплексностью факторов, послуживших их образованию. Надо сказать, что исследованиями жизненных форм занимался еще А. Гумбольдт, а сам термин ввел Е. Варминг в 1844 г., пока не удалось создать более или менее общую классификацию жизненных форм в соотношении их с экологическими типами.
В целом же дополнительное разнообразие, которое вносится в биоценозы за счет создания их "пищевой" структуры, в еще большей степени повышает устойчивость экосистем. Чем сложнее условия, тем больше количество отличающихся по биологическим особенностям видов могут освоить данный биотоп. С возрастанием биологической сложности состава экосистем уменьшается объем экологических ниш. По А. Тинеману, в благоприятных условиях высоко число видов, но каждый из них представлен относительно небольшим числом особей; в неблагоприятных условиях это соотношение меняется на обратное, это так называемое правило "числа видов и числа особей".
163
И.А. Шилов (2000) приводит меткое выражение известного биолога А. Уоллеса о том, что в тропических лесах легче за день поймать по одной бабочке ста видов, чем сто бабочек одного вида. "Пищевая" структура биоценозов создает определенную свободу в формировании сообщества: биологически сходные виды могут замещать друг друга в разных экосистемах.
Жизненная форма растений по К. Раункнеру (1903):
- фанерофиты (от греч. "видимый" и "растений")-растения, у которых почки возобновления находятся на некотором расстоянии от поверхности почвы (выше 25 см); это главным образом деревья и крупные кустарники;
- хамефиты (от греч. "на земле") - растения (мелкие кустарники, кустарники, низкорослые суккуленты), у которых почки возобновления расположены низко над поверхностью почвы (ниже 25 см);
- гемикриптофиты (от греч. "полу" и "скрытый") - многолетние травянистые растения, у которых почки возобновления располагаются на уровне почвы и защищены отмершими листьями или снегом;
- криптофиты - многолетние травы, у которых почки возобновления закладываются в луковицах, клубнях, корневищах и находятся в почве или под водой, благодаря чему они защищены от прямого воздействия среды;
- терофиты (от греч. "лето") - однолетние травянистые растения, которые переживают неблагоприятный для их жизнедеятельности период в виде семян.
Жизненные формы животных по А.Н. Формозову: наземные; подземные (землерои); древесные; воздушные; водные.
В процессе взаимоотношений и взаимодействий видов и отдельных живых организмов происходит их естественный отбор, приспособительная изменчивость. Вопросам естественного отбора мы уделим особое внимание несколько ниже.
Факторы среды как по отдельности, так и в комплексе при воздействии на живые организмы заставляют их изменяться, адаптироваться к этим факторам. Эта способность носит название экологической валентности или пластичности. Пластичность, или экологическая валентность, каждого вида различна и по-разному сказывается на способности живых организмов выживать в условиях меняющихся факторов среды. Если к биотическим факторам организмы не только приспосабливаются, но и могут на них воздействовать, изменяя другие живые организмы, то с абиотическими факторами среды это невозможно: организм может к ним приспособиться, но не в состоянии оказать на них сколько-нибудь значимое обратное влияние.
Существует два типа приспособления к внешним факторам. Первый, по И.А. Шилову (2000), заключается в возникновении определенной степени устойчивости к данному фактору, способности сохранять функции при изменении силы его воздействия. Это пассивный путь
164
адаптации - адаптация по принципу выносливости (толерантности). Такой тип приспособления формируется как характерное видовое свойство и реализуется преимущественно на клеточно-тканевом уровне. Второй тип приспособления - активный. В этом случае организм с помощью имеющихся у него специфических адаптивных механизмов компенсирует изменения, вызванные воздействующим фактором, при этом внутренняя среда организма остается относительно постоянной. Этот вид адаптации поддерживает динамическое равновесие внутренней среды организма.
Выделяют организмы, приспособленные к широкому диапазону имеющихся факторов среды-эврибионты или эвритопные, и организмы, меняющиеся только в узком диапазоне - стенобионты или стенотопные, соответственно, например, для приспособленных к жизни в широком температурном диапазоне - эвритермные, в узком - стенотермные.
Адаптации вырабатываются у живых организмов под влиянием множества динамичных экологических факторов, зависящих от геологических, климатических и даже астрономических процессов. Адаптации должны быть закреплены в наследственной информации, что достигается в результате эволюции, которая должна быть синхронизирована как со сроками воздействия факторов, так и со временем существования живого организма и вида.
Адаптации могут быть морфологическими, выраженными в приспособлении строения (формы) организмов к факторам среды, примером могут служить различия в размерах ушных раковин у лесных и степных ежей; физиологическими - приспособление пищеварительного тракта к составу пищи, примером является строение желудка с наличием дополнительного отдела у жвачных травоядных; поведенческими или экологическими - приспособление поведения животных к температурным условиям, влажности и т. д., примером может служить зимняя спячка у ряда животных: грызунов, медведей и др.
Приспособления к питанию растениями обнаруживаются, как было отмечено, и на физиологическом уровне. Особенно выражены они у животных, питающихся грубыми тканями вегетативных частей растений, содержащими большое количество клетчатки. В организме большинства животных не продуцируются целлюлозолитические ферменты, а расщепление клетчатки осуществляется симбиотическими бактериями и некоторыми простейшими кишечного тракта.
Среди млекопитающих переработка клетчатки особенно специализирована у жвачных копытных, желудок которых имеет сложное строение (рис. 43). Наиболее объемистый его отдел - рубец - служит вместилищем, в котором "проглоченный" корм перемешивается со слюной и подвергается воздействию ферментов симбиотических бактерий и простейших. Продуценты бактериологического сбраживания клетчатки (в основном летучие жирные кислоты - уксусная, янтарная,
165

Рис. 43. Строение желудка жвачных копытных (по К. Шмидт-Нильсену, 1982):
1,3 - рубец;
2 - тонкая кишка;
4 - пищевод;
5 - сетка;
б - книжка;
7 - сычуг
масляная) всасываются здесь же, в рубце. Оставшаяся часть растительной массы после вторичного пережевывания подвергается действию желудочного сока в другом отделе желудка - сычуге - и далее следует обычным путем через кишечник, последовательно обрабатываемая различными ферментами. При таком типе питания в кишечник наряду с растительной массой попадает большое количество бактерий и простейших, клетки которых служат источником белкового питания; кроме того, некоторые бактерии в рубце способны синтезировать белок из аммонийных солей и мочевины.
У других млекопитающих основным местом переработки клетчатки является толстая кишка, особенно ее слепой вырост. Химизм этих процессов сходен с пищеварением в рубце. У зайцеобразных и многих грызунов в связи с таким типом пищеварения выделяется два типа помета: первичный, который выводится из организма, и вторичный. Поедание первичного помета способствует утилизации содержащегося в нем белка (включая и бактериальные белки), а может быть, и "подсеву" регулярно теряемой с экскрементами кишечной флоры. Аналогичным образом в области толстого кишечника и его слепых выростов идет переваривание клетчатки у питающихся растительными тканями птиц (куриные, гусеобразные, врановые и др.).
Одним из видов адаптации является наличие приспособительной окраски у ряда животных как средство пассивной защиты. Многие животные средних и высоких широт зимой имеют белую окраску - заяц, горностай, песец; а скрывающая (расчленяющая) окраска, например чередование светлых и темных полос или в виде пятен у тигра, зебры, леопарда приводит к тому, что они плохо видны на открытой местности или в меняющемся свете в лесу, в тайге.
В качестве же наиболее известного вида адаптации стоит остановиться на мимикрии - подражании животных и растений определенным предметам как живой, так и неживой природы. В 1862 г. английский натуралист Г. Бейтс при изучении животных в бассейне р. Амазонки установил подражание незащищенного съедобного вида (имитатора) другому, неродственному ему, который невкусен и опасен
166
для хищника ("мимикрия Бейтса"). Так, многие виды безвредных змей сходны по внешнему виду с ядовитыми и др. Такие имитаторы широко встречаются среди насекомых, амфибий, птиц, рептилий. В 1878 г. немецкий зоолог Ф. Мюллер при изучении насекомых в Бразилии также показал, что различные несъедобные виды животных имеют сходную предупреждающую окраску, которые всем им приносит пользу, так как хищники, часто сталкиваясь с несколькими видами - имитаторами, привыкают избегать тех, кто входит в "кольцо мимикрии" (В.А. Вронский, 1996).
Адаптация к любому фактору связана с затратами энергии. В зоне оптимальных условий для жизнедеятельности организмов адаптивные механизмы отключены, и энергия расходуется только на фундаментальные жизненные процессы, это так называемые энергозатраты на базальный метаболизм. Если значения фактора воздействия выходят за пределы оптимума, то организм или вид включает адаптивные механизмы, функционирование которых сопряжено со значительными энергозатратами, тем большими, чем ближе смещение в зону стресса. При этом увеличение энергетических расходов резко ограничивает возможный набор форм жизнедеятельности организма: чем дальше от оптимума находится количественное выражение фактора, тем больше энергии направленно расходуется на адаптацию и тем меньше "степеней свободы" в проявлении иных форм деятельности (И.А. Шилов, 2000).
Взаимодействие биотических и абиотических факторов выражается комплексно, хотя влияние одного или другого из них - может быть и раздельным, большей или меньшей интенсивности. Совокупное действие на организм нескольких факторов среды обозначают термином констелляция. Экологически важно то обстоятельство (по И.А. Шилову, 2000), что констелляция не представляет собой простой суммы влияния факторов; при комплексном воздействии между отдельными факторами устанавливаются особые взаимодействия, когда влияние одного фактора в какой-то мере изменяет (усиливает, ослабляет и т. п.) характер воздействия другого. В качестве примера можно привести факт различий в реакции газообмена у рыб в условиях разной солености воды.
Принципиальных путей адаптации к определенному фактору немного: адаптивные механизмы полностью запрограммированы физико-химической природой данного фактора. В ходе эволюции жизни на Земле адаптация к лимитирующим факторам нередко определяла наиболее фундаментальные изменения в морфологии и физиологии.
"Так, выход позвоночных животных на сушу был невозможен без применения двух принципиальных лимитирующих факторов: малой плотности среды и низкой ее влажности. В водной среде, плотность которой сопоставима с плотностью тела животных, организмы оказывались "парящими" в воде, локомоторная система функционировала
167
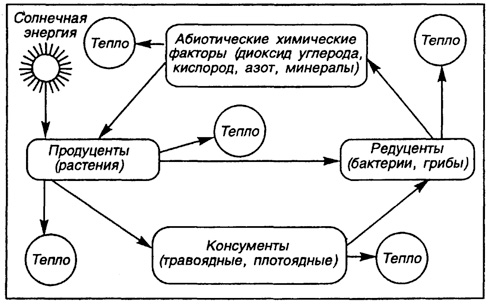
Рис. 44. Общая схема пищевой цепи
лишь для придания телу поступательного движения. В воздушной среде такой принцип локолюции1 оказался непригодным: благодаря малой плотности воздуха наземные животные прижаты к субстрату весом собственного тела. Эволюционно эта задача решалась путем формирования конечностей рычажного типа, способных одновременно обеспечить функцию опоры на субстрат и функцию поступательного движения. "Четвероногие", т. е. наземные позвоночные, таким образом, возникли как результат приспособления к малой плотности среды.
Низкая влажность воздушной среды лимитировала функционирование водного типа дыхательной системы, поскольку создавала постоянную угрозу высыхания поверхности дыхательного эпителия. Приспособление к дыханию в новой среде - появление легких; у легочно-дышащих животных дыхательная поверхность не соприкасается с воздушной средой, а связана с ней узкими воздухоносными путями и снабжена системой желез, увлажняющих как поверхность дыхательного эпителия, так и подводимый к нему воздух. Одновременно шли эволюционные перестройки строения покровов, направленные на снижение потерь влаги через поверхность тела" (Шилов И.А., 2000, с. 225).
В конкретных биоценозах все живые организмы представлены популяциями многих видов, состав которых специфичен для каждого
168

Рис. 45. Пастбищная сеть питания в наземном биоценозе
169

Рис. 46. Детритная пищевая сеть
конкретного сообщества. Взаимоотношения между видами, находящимися на различных функционально связанных пищевых (трофических) уровнях, образуют систему трофических (пищевых) цепей, которые в свою очередь формируют трофическую (пищевую) структуру биоценоза (рис. 44).
Трофические (пищевые) цепи и трофические уровни, как было уже показано выше, имеют важнейшее значение для функционирования биогеоценозов и играют роль определяющего фактора во взаимоотношениях живых организмов. При изучении любой биотической системы можно проследить бесчисленное множество путей движения вещества, запасенной энергии и информации, можно установить, что один организм поедается другим, тот - третьим и т. д., т. е. образуется трофическая цепь. При этом выясняется, что трофические цепи почти никогда не бывают изолированными. Известным фактом является то, что травоядные животные поедают не одно растение, а питаются сразу несколькими видами, а сами могут служить пищей нескольким видам консументов. Практически все пищевые цепи соединены между
170
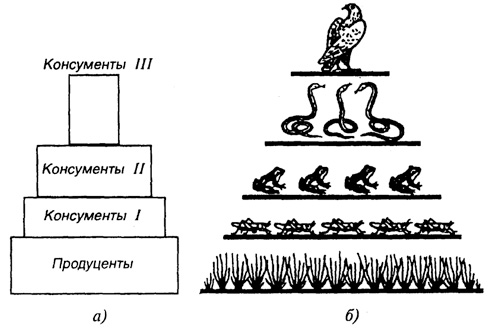
Рис. 47. Упрощенная схема экологической пирамиды
(а) и пирамиды чисел
(б)
собой и образуют сложную сеть пищевых взаимоотношений (рис. 45, 46).
Все продуценты относятся к первому трофическому уровню, все первичные консументы, независимо от того, живыми или мертвыми продуцентами они питаются, относятся ко второму трофическому уровню, соответственно консументы 2-го порядка - к третьему и т. д. Как правило, количество трофических уровней не превышает трех-четырех. Б. Небел (1993) этот вывод подтверждает следующим: общую массу организмов (их биомассу) на каждом трофическом уровне можно подсчитать путем сбора (или отлова) и последующего взвешивания соответствующих выборок растений и животных. Таким образом установлено, что на каждом трофическом уровне биомасса на 90-99 % меньше, чем на предыдущем. Из этого нетрудно представить, что существование большого числа трофических уровней невозможно из-за того, что биомасса весьма быстро приблизится к нулю. Графически это представляется в виде пирамиды биомассы (рис. 47).
По Ю. Одуму, "экологическая пирамида, представляющая собой трофическую структуру, основанием которой служит уровень продуцентов, а последующие уровни образуют этажи и вершину пирамиды, может быть трех основных типов: 1) пирамида чисел, отражающая численность отдельных организмов; 2) пирамида биомассы, характеризующая общий сухой вес, калорийность или другую меру общего количества живого вещества; 3) пирамида энергии, показывающая величину потока энергии и (или) "продуктивность" на последовательных трофических уровнях".
Снижение биомассы по трофическим уровням объясняется тем,
171
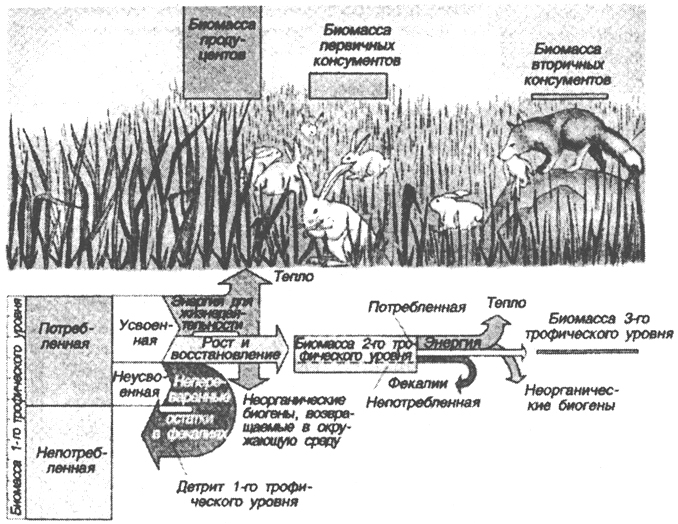
Рис. 48. Снижение биомассы с повышением трофического - уровня (по Б. Небелу, 1992)
что на более высоком трофическом уровне большая часть потребляемой в пищу боимассы идет не на формирование тела организмов, а на получение из этой пищи запасенной в ней энергии (рис. 48).
Еще раз нелишним будет подчеркнуть значимость продуцентов для поддержания функционирования экосистем: без производимой биомассы экосистема довольно быстро "съела бы" саму себя.
Биотические факторы проявляются в коакциях организма при совместном обитании и имеют весьма разнообразный и можно сказать даже причудливый характер. Виды взаимодействий подразделяются следующим образом.
- Нейтральные, или независимые, отношения устанавливаются довольно часто между разными видами, которые совместно обитают в пределах одного биогеоценоза, на одной территории. В данном случае одни организмы не оказывают влияния на другие непосредственным образом, хотя имеют общие потребности в пище, но не вступают во взаимоотношения друг с другом, например олени или ложи живут в одном лесу с белками, тетеревами, но не мешают друг другу, хотя питаются растительной пищей и даже из одного растительного яруса.
172
Разные виды копытных в саваннах Африки образуют на пастбищах совместно пасущиеся стада, но не мешают друг другу и питаются растительной пищей из разных растительных ярусов.
- Конкуренция - это тип взаимоотношений, который обычно имеет место у близких видов (как правило, не более двух), имеющих сходные потребности и обитающих на одной территории. Растения, которые растут рядом, вынуждены соперничать в борьбе за воду, солнечный свет, биогены, да и за жизненное пространство. Присутствие одного вида или организма уменьшает ресурсы пищи, сокращает возможность для расселения другого вида и организма. В качестве примеров можно привести конкуренцию различных видов грызунов, обитающих на одном поле, или конкуренцию злаков и сорняков на том же поле, волков и лис, живущих в одном лесу. В результате такого рода взаимодействия более слабый конкурирующий организм или вид вытесняется более сильным, а в худшем случае погибает.
Конкуренция возникает в некоторых случаях и внутри одного вида между отдельными организмами, что в большинстве случаев вызвано тем, что эти организмы используют одни и те же ресурсы, и протекает тем острее, чем ограниченнее эти ресурсы. К такому виду конкуренции относятся, например, борьба за лучшее место гнездования, за самку для продолжения рода. В некоторых случаях за счет адаптации к условиям среды, например у некоторых животных - бабочек, амфибии-личинки и взрослые особи питаются разной пищей и живут в разных ярусах экосистемы. Это в целом снижает давление конкуренции. Иными cловами, возможность организмов или видов меняться при воздействии факторов среды и наличие ресурсов позволяет уменьшать конкурентную борьбу, в частности, за счет того, что разные виды животных адаптированы к питанию неодинаковой пищей, в разных местах и (или) в разное время.
Хищничество. Организмы, которых относят к хищникам, питаются другими организмами, уничтожая свою жертву. Таким образом, среди живых организмов следует выделять еще одну классификационную систему, а именно "хищники" и "жертвы". Отношения между такими организмами складывались на протяжении всей эволюции жизни на нашей планете. Организмы-хищники выступают в роли природных регуляторов численности организмов-жертв. Увеличение численности "хищников" приводит к уменьшению численности "жертв", это, в свою очередь, снижает запасы пищи ("жертв") для "хищников", что в целом диктует снижение численности "жертв" и т. д. Таким образом, в биоценозе постоянно происходят колебания численности хищников и жертв, в целом же устанавливается определенное равновесие на какой-то период времени в пределах достаточно устойчивых условий среды.
Следует отметить, что в условиях конкурентной борьбы происходит изменение даже в поведении хищников и жертв, как это происходит
173
в стадах оленей, когда они выстраиваются в круг для отражения нападения волков повернутыми в сторону нападающих рогами. Довольно часто хищники в первую очередь уничтожают слабые и больные организмы, что является положительным фактором в поддержании здоровья живых организмов. Если поедают друг друга особи одного вида, то такого рода отношения носят название каннибализма. Взаимоотношения типа хищничества не являются присущим исключительно животным; среди растений известны насекомоядные - росянки, среди микроорганизмов - явление фагоцитоза.
Кроме пищевых взаимоотношений между отдельными организмами и видами возможны и иные, например паразитизм, симбиоз, "нахлебничество".
Паразитизм представляет собой отношения, при которых один вид живет за счет ресурсов другого. Под ресурсами в данном случае понимается, что один организм служит для другого и источником пищи, и местом обитания. Самым простым примером являются гельминты - паразитические черви, обитающие в пищеварительных и выделительных системах животных, да и человека. В целом же к паразитам чаще всего относят простейших, бактерии, грибы, червей. В отличие от хищника паразит не убивает свою жертву, а достаточно длительное время живет за счет ее ресурсов. Паразитизм может быть временным и постоянным, иногда в процессе жизни возможна смена хозяев. В процессе эволюции у паразитов выработались приспособления к осуществлению своей жизнедеятельности - упрощение органов, редукция частей тела. Например, у паразитических червей полностью редуцированы органы пищеварения, а растения-паразиты теряют значительную часть хлорофилла.
Мутуалистические отношения или мутуализм - это один из способов реализации пищевых цепей. В целом в пищевых цепях подразумевается, что один из видов извлекает выгоду, а другому наносится вред. Однако в природе существует немало случаев, когда виды вступают во взаимовыгодные отношения,- этот феномен и носит название мутуализма. Классическим примером являются лишайники, которые собственно и представляют собой не один, а два организма - гриб и водоросль. Гриб обеспечивает водоросли защиту, позволяя ей выжить в таких условиях малой влажности, где она собственно сама выжить не может, ну а водоросль, как продуцент, поставляет грибу пищевые ресурсы. Кстати и сами грибы сосуществуют с корнями деревьев, где процессы положительного мутуализма или симбиоза аналогичны лишайникам; можно также вспомнить отношения актинии и рака-отшельника, цветков растений и насекомых и т. д.
К неконкурентным отношениям организмов относится "нахлебничество" - это взаимоотношения, когда один вид извлекает пользу от присутствия другого, которому большей частью наличие этого вида безразлично: рыбы-лоцманы и рыбы-прилипалы следуют за акулами
174
и др. В отдельных случаях отмечен также такой вид отношений, который можно назвать квартиранством, например в полости рта голотурии живут мелкие морские организмы; мхи, лианы и другие растения-эпифиты только прикрепляются к стволам и ветвям деревьев для лучшего доступа к воздуху, влаге и свету, но существуют за счет своего собственного фотосинтеза.
Таким образом, среди организмов, населяющих экосистему (биогеоценоз), существуют весьма непростые виды взаимоотношений, как пищевые, так и непищевые, что в силу разнообразия их проявления повышает устойчивость жизненных процессов.
Вид.Каждый вид живых организмов обладает наиболее существенным геохимическим свойством, а именно специфичностью обмена веществ с внешней средой. В целом вид является формой существования жизни. Вследствие специфичности в метаболизме только разнообразие видов в составе каждого трофического уровня, а следовательно, и в составе экосистемы обеспечивает максимальную эффективность использования источников и форм энергии для синтеза первичной продукции и трансформации вещества на разных этапах биогенного круговорота, вплоть до полной минерализации и повторного вовлечения в цикл. Кроме того, многообразие в биогеоценозе равнозначных видов по функциональным признакам является механизмом обеспечения устойчивости собственно круговорота, т. е. потоков вещества, энергии и информации по трофическим цепям. Кстати, природа разработала специальный механизм замены "выпавшего" вида в трофической структуре биогеоценоза другим равнозначным по функциональным возможностям видом, который при нормальных условиях функционирует в том же биогеоценозе. Биологическое разнообразие или разнокачественность жизни как фактор устойчивости биологических круговоротов на уровне биогеоценоза реализуется путем увеличения видового разнообразия. Биоразнообразие в настоящее время признается как один из глобальных факторов, определяющих саму возможность существования жизни как планетарного явления. Неслучайно вопросу сохранения биоразнообразия уделено такое большое внимание в "Повестке дня на XXI век", принятой на глобальном экофоруме в Рио-де-Жанейро в 1992 г.
Популяция. Виды живых организмов обеспечивают устойчивое поддержание биогенного круговорота в биогеоценозе на уровне популяции. Изучением популяций занимаются достаточно давно, и к настоящему времени сложилось определенное понимание особенностей их функционирования. Под популяцией понимают исторически сложившуюся естественную общность особей живых организмов одного вида, связанных генетически, населяющих общие места обитания и реализующих закономерные функциональные взаимодействия.
По A.M. Гилярову (1990, с. 38): "популяция - это любая, способная к самовоспроизведению, совокупность особей одного вида, более или
175
менее изолированная в пространстве и времени от других аналогичных совокупностей того же вида". Особи в популяции скрещиваются друг с другом и таким образом размножаются. Без этого условия вид из экосистемы исчезнет.
В современной популяционной экологии эту общность организмов рассматривают как биологическую систему надорганизменного уровня, обладающую собственными функциями и структурно обусловленную (С.С. Шварц, 1980, И.А. Шилов, 2000).
Рассмотрение популяций обычно проводят с двух главных позиций. Во-первых, популяция является формой жизни конкретного вида живых организмов в условиях среды их обитания. В этом случае осуществляется важнейшая функция популяции - обеспечение выживания живых организмов составляющего ее вида и воспроизведение вида в данных условиях. Данная функция обеспечивается ориентированными адаптациями (приспособительными изменениями) отдельных особей вида, имеющих морфологическую общность, а также сформированными закономерными взаимоотношениями. Все это поддерживает и регулирует возможности размножения (воспроизводство) особей вида. Процесс этот носит непрерывный характер, выражающийся в смене отдельных индивидов внутри популяции, и поэтому при оптимальных условиях жизнедеятельности существование популяции может продолжаться неограниченно долго - популяция бессмертна как структурная единица.
Во-вторых, популяция, являясь структурной единицей биогеоценоза (экосистемы), выполняет одну из главнейших его функций, а именно участвует в биологическом круговороте. В данном случае реализуется видоспецифическая особенность типа обмена веществ. Популяция представляет вид в экосистеме и все межвидовые взаимоотношения осуществляются в ней на популяционном уровне. Устойчивая реализация функции участия в биогенных процессах определяется специфическими механизмами авторегуляции, которые создают условия самоподдержания популяции как системы в изменяющихся внутренних и внешних факторах среды обитания.
В целом популяции обладают всеми признаками самостоятельной функционирующей биологической системы. Популяция имеет пространственное расположение входящих в нее элементов, они структурированы, что позволяет оптимально использовать ресурсы среды, а также создает условия для бесперебойного осуществления внутрипопуляционных взаимоотношений.
Однако особи, входящие в вид, формирующий данную популяцию, не являются абсолютно идентичными в осуществлении внутрипопуляционных взаимоотношений. Эти взаимоотношения образуют определенную функциональную систему. Даже при условии общего морфофизиологического типа (сходстве) отдельные особи неравноценны
176
по своему участию в жизнедеятельности популяции. Таким образом, популяция структурирована еще и функционально.
Особи в популяциях находятся в состоянии постоянного обмена информацией, что представляет собой специфический механизм взаимодействия живых организмов. На базе генетической разнокачественности организмов, а у высших животных на поведенческом уровне в популяции вырабатываются весьма сложные, но очень эффективные механизмы авторегуляции.
Следует отметить чрезвычайно важную особенность функционирования популяции: взаимодействие особей со средой осуществляется через физиологические реакции, причем эти реакции исключительно индивидуальны, но направленность их такова, что в целом они реализуют общепопуляционные функции. Как утверждает И.А. Шилов (2000, с. 46), "физиология отдельных организмов в составе популяции как бы решает двойную задачу: физиологические процессы обеспечивают, с одной стороны, жизнь и адаптацию самой особи, а с другой - устойчивое поддержание функций целостной популяции".
Организация на популяционном уровне связана главным образом с регулированием численности и плотности популяций. Численность популяции - это общее число особей, обитающих на какой-либо территории или в сообществе. Плотность популяции - это величина, определяемая числом особей или биомассой по отношению к единице пространства (территории или даже сообществу, если его можно описать пространственно). Пространственная структура популяции определяется особенностями расселения популяции на территории. Часто отдельные особи образуют семьи, стаи, стада и т. п. С помощью специальных сигналов они отмечают занимаемый участок (пение птиц, пахучие вещества или экскременты у животных и т. д.). Отмечены даже определенные маршруты миграции. При резком возрастании численности популяции возможно изменение пространственной структуры за счет конкурентной борьбы.
В естественных условиях численность и плотность популяции определяются регулирующими (управляющими) экологическими факторами. Объем популяции определяется стациальной емкостью экосистемы (стация - место обитания). Размеры популяции небезграничны; каждая популяция имеет верхний и нижний пределы. Способность популяции к увеличению характеризуется рождаемостью. Абсолютная (максимальная или физиологическая) рождаемость - это теоретически возможное количество особей в идеальных условиях, когда размножение ограничивается только физиологическими факторами (для каждой данной популяции это величина постоянная). Кроме этого различают экологическую или реализуемую рождаемость. Рождаемость определяется способностью популяции воспроизводить новые особи с определенной частотой (число детенышей, отложенных яиц, икринок
177
у животных, семян и спор у растений). У микроорганизмов рождаемость зависит от скорости деления клеток.
Смертность характеризуется числом особей, умерших за определенный период, т. е. скоростью уменьшения численности популяции. На разных стадиях развития гибель особей существенно различна. Смертность рыб на стадии икринок и мальков значительно выше, чем среди взрослых особей. Чем выше развит инстинкт заботы о потомстве, как у высших животных, тем больше особей выживает в раннем возрасте. В противном случае рост смертности компенсируется высокой плодовитостью особей.
Каждая популяция структурирована по возрасту, численности, пространству, полу. Возрастная структура популяции определяется соотношением особей разного возраста; при этом установлено, что это соотношение непостоянно, а имеет довольно сложные колебания. В стабильной популяции рождаемость равна смертности и численность популяции почти не меняется, разновозрастные группы находятся примерно в одинаковом соотношении. В растущих популяциях рождаемость превышает смертность и численность увеличивается. Половая (сексуальная) структура определяется соотношением полов, количеством самцов и самок в популяции. Практически нет популяций, сходных по этой структурной характеристике (у животных известны "гаремные" и "парообразующие" формы).
По отношению к популяции выделяют три возраста - пререпродуктивный, репродуктивный, пострепродуктивный. Существует константа стабильного распределения по возрастам. Для мелких организмов характерен короткий жизненный цикл, для крупных - более длинный. Как отмечается многими специалистами, в популяции имеет место компенсаторный механизм, который диктует следующую закономерность: высокое выживание обусловливает высокую вероятность снижения выживания в последующие годы. Распределение особей может быть случайным, если среда существования популяции однородна и особям нет необходимости объединения в группы и т. п.; равномерным - при наличии сильной конкуренции между особями, что приводит к их распределению по пространству; групповым - в виде стаи, стад и т. п., если это определяет оптимальность выживания.
Параметры популяции весьма изменчивы во времени и зависят от множества факторов. В популяции идут два противоположных процесса - изоляции и агрегации. Факторы изоляции - конкуренция между особями за пищу при ее недостатке и прямой антагонизм. Это ведет к равномерному или случайному распределению особей. Выше мы уже рассматривали вопрос о конкуренции. Для агрегации характерны два последствия: увеличение внутривидовой конкуренции и взаимопомощи, способствующей выживанию группы в целом. "У особей, объединенных в группу, по сравнению с одиночными особями нередко наблюдается снижение смертности в неблагоприятные периоды или
178
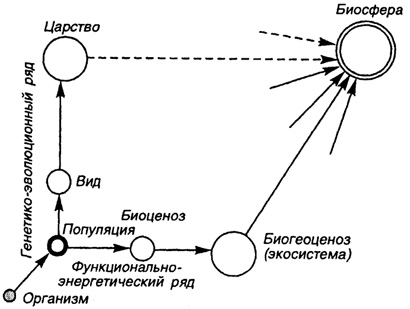
Рис. 49. Положение популяции в структуре биологических систем в биосфере
(по И.А. Шилову, 1988)
при нападении других организмов, поскольку в группе поверхность их соприкосновения со средой по отношению к массе меньше и поскольку группа способна изменять микроклимат или микросреду в благоприятном для себя направлении" (Ю. Одум, 1975, с. 269).
Периодический уход и возвращение на данную территорию называют миграцией. Изменения численности организмов во времени называют динамикой популяции. Периодические колебания называются осцилляциями, непериодические - флуктуациями. Первые связаны с регуляторными изменениями факторов среды, сезонными ритмами. Иногда отмечаются вспышки численности особей незакономерного характера, флуктуационного типа, которые называются попу-ляционными волнами.
Способность популяции к авторегуляции носит название гомеостаза популяции. Обычно популяция находится именно в этом состоянии - динамического равновесия, которое достигается за счет чередования положительных и отрицательных обратных связей. При росте численности особей сокращаются запасы пищи, что влечет уменьшение численности за счет увеличения смертности, а сократившаяся популяция позволяет накопить запасы пищи и ведет к увеличению рождаемости, т. е. росту численности и т. д.
Таким образом, структурированность, интегрированность составных частей (особей) или целостность, авторегуляторные механизмы и способность к адаптациям - это в целом определяет популяцию как биологическую систему надорганизменного уровня (рис. 49).
Организм. Главной составляющей популяции является особь или организм. Организм представляет собой структурно-функционально организованную подсистему. Она занимает вполне определенное положение в популяционных взаимосвязях и выполняет четко обусловленные функции в общепопуляционных процессах. Именно организм представляет собой ту биологическую единицу, на уровне которой осуществляется обмен веществ и которая собственно находится в тесных взаимосвязях как с другими организмами и более крупными биологическими системами, так и с факторами неживой природы.
179
Еще в конце XIX в. К. Бернар рассматривал организм как биологический объект, как систему интегрированных обособленных морфологических частей. Он утверждал, что за счет стабильности физико-химических условий во внутренней среде организма реализуются свобода и независимость живых организмов в изменчивых условиях среды.
Значительным достижением в биологической науке явилось установление в 1929 г. У. Кенноном гомеостаза (от греч. "тоже" и "состояние") организма, который отражает его целостность и возможность поддерживания постоянства внутренней среды. В 1949 г. это было серьезно дополнено и обосновано в работе П.К. Анохина, в его концепции функциональных физиологических систем.
По И.А. Шилову (2000, с. 47), "функция обмена веществ в организме определяется согласованной деятельностью различных систем органов; регуляция метаболических процессов лежит в основе адаптации жизнедеятельности к изменчивым условиям среды. Устойчивость обменной функции в глобальном масштабе определена способностью живых организмов к самовоспроизведению - уникальной функцией живого вещества". Следует отметить, что в реализации этой функции кроме вещественного круговорота и регуляции потоков энергии огромная роль принадлежит информационным взаимодействиям и во многом самовоспроизведение является реализацией информационных функций организмов. Рассмотрение живого организма (особи) как носителя определенного информационного объема еще далеко не завершено и требует дополнительных исследований. Это имеет значение и для понимания закономерностей функционирования особей внутри популяций, а также и на межвидовом уровне.
Чрезвычайно важным представляется рассмотрение организма как среды жизни. Весь комплекс физиологических процессов на уровне организма подразделяется на реализующиеся во внутреннем функционировании и на "реакционные" (ответные) на воздействие окружающих факторов среды. Первая группа реакций - это составляющие сущности жизни: поглощение, переваривание и усвоение пищи, клеточный обмен веществ (собственно метаболизм), дыхание, водно-солевой обмен и др. Суммарно они создают условия для жизни организма, т. е. если они есть в наличии, то организм можно отнести к "живому". Если же рассмотреть эти процессы глобально, то опять же суммарно они создают возможность функционирования соответствующего данному виду обмена в трофической системе или биогенном круговороте.
Такое функционирование встречает достаточно активное противодействие со стороны окружающей организм среды, которая воздействует не только многофакторно, но и весьма динамично. Вследствие этого вторая группа физиологических реакций направлена на адаптацию
180
организма к действию факторов, влияющих на протекание жизненно важных процессов в организме, на функционирование первой группы процессов. Проще говоря, эта вторая группа относится к процессам "выживания" в сложной и изменчивой среде. Интегрированный результат данных физиологических процессов выражается в поддержании гомеостаза организма, в создании относительного постоянства условий его внутренней среды.
Кроме вышеуказанных "общеорганизменных" функций наличие гомеостаза организма существует еще одна очень важная особенность: живое вещество как бы создает еще одну среду обитания, а именно возможность заселения организма другими живыми существами для постоянного или временного обитания. Это созданная жизнью новая биотическая среда обитания. К существам, которые заселяют эту среду, многие специалисты относят вирусы. Так, ИЛ. Шилов (2000) считает, что исключительная простота их устройства является вторичным явлением, даже скорее это вновь возникшая форма живых существ, полностью освоившая внутриклеточную среду в организмах других уровней. Вторым подтверждением этого тезиса является то, что вирусы обладают высокой степенью сложности и разнообразия генетической системы. Упрощение строения, ставшее возможным благодаря обязательным безусловным связям вирусов с хозяином-организмом, обеспечивающим стабильные условия жизни, затронуло даже фундаментальные свойства, присущие подавляющему большинству форм жизни: вирусы не обладают раздражимостью и лишены собственного аппарата синтеза белка. Вирусы не способны к самостоятельному существованию, и их связь с клеткой - это не только пространственная, но и жесткая функциональная связь, с которой клетка и вирус представляют некое единство.
Выше мы уже останавливались на том, что существует группа живых организмов, которая использует другие как среду обитания, это так называемые паразиты. В некоторой степени к такому взаимодействию можно отнести различные формы симбиоза. Во всех случаях возникает тесно обусловленная функциональная связь между организмами, а созданные взаимоотношения развиваются коадаптационно.
Наличие вновь созданной биотической среды обитания и вторичных форм жизни в живых организмах приводит к выводу, что целостность биосферы заключена не только в ее экосистемной структурированности на глобальном уровне, но и в том, что организмы - это первое звено в циркуляции вещества в биологических системах разного уровня, функционирует как специфическая среда, в которой в свою очередь формируются и функционируют достаточно богатые сообщества живых организмов.
181
1
Локолюция (от лат.
lokus - место и
motto - движение), движения животных и человека, обеспечивающие активное перемещение в пространстве; важнейшее приспособление к обитанию в разнообразных условиях среды. Типы локолюции в процессе эволюции животных менялись и усложнялись (СЭС, 1980, с. 732).
