Полимерные цепи (вне зависимости от регулярности их строения) под влиянием теплового движения и межмолекулярного взаимодействия ассоциируются во флуктуационные, более или менее упорядоченные пачки. Пачки под влиянием теплового движения то воссоздаются, то распадаются. Однако вследствие больших размеров макромолекул время жизни пачек может быть весьма большим. Если продолжительность жизни роя молекул низкомолекулярной жидкости составляет 10-8 с и менее, то в случае твердого полимера она возрастает до многих лет. Чем менее гибки макромолекулы, тем больше время жизни пачек.
Контакт между пачками может осуществляться как в результате межмолекулярного взаимодействия цепей, расположенных во внешних слоях пачки, так и за счет так называемых проходных
153

Рис. 3.17. Схематическое изображение проходных цепей
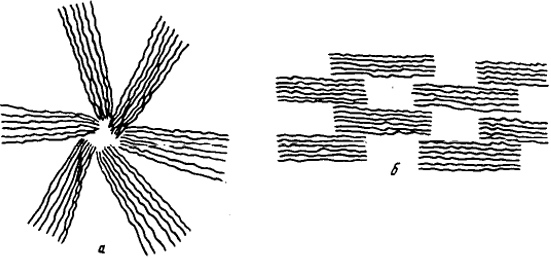
Рис. 3.18. Схематическое изображение надмолекулярных структур:
а - сферолитной; б - фибриллярной
цепей. В последнем случае одна часть макромолекул находится в одной пачке, а другая - в другой (рис. 3.17). Пачки макромолекул в свою очередь агрегируются в более крупные надмолекулярные образования - микрофибриллы.
Если полимер построен из регулярных полимерных цепей, т.е. способен к кристаллизации, то плотность упаковки макромолекул в различных частях пачек различна: в аморфных областях меньше, в кристаллических - больше. В свою очередь пачки макромолекул ассоциируются в микрофибриллы так, что между пачками появляются области с малой плотностью упаковки или даже пустоты. Дальнейшее усложнение надмолекулярной организации
154
полимеров идет по пути образования либо сферолитных, либо фибриллярных структур (рис. 3.18). Сферолитные структуры возникают преимущественно в изотропных средах. Приложение силового поля способствует образованию преимущественно фибриллярных структур.
Рассмотрим, например, структуру волокна на основе природной целлюлозы. Рентгенографическими и ИК-спектроскопическими исследованиями установлено, что элементарное звено целлюлозы - ангидро-β-D-глюкоза - имеет конфигурацию "кресла", а полимерная цепь построена из регулярно соединенных между собой, правильно расположенных в пространстве ангидроглюкозных звеньев:

Природная целлюлоза - стереорегулярный синдиотактический полимер. Схематически макромолекулы целлюлозы представляют собой слегка свернутую спираль - "ленту" - сечением 0,39 × 0,83 нм. Агрегация молекул целлюлозы обусловлена большим числом полярных гидроксильных групп и регулярным строением полимерных цепей.
Ленты укладываются более или менее параллельно в одной плоскости по 10-15 штук, образуя пачки молекул из 8-10 слоев (рис. 3.19). Вследствие относительной жесткости макромолекул и прочности межмолекулярных связей такие пачки существуют практически бесконечно долго. В результате образуется пачка, сечение которой составляет примерно 5 × 8 нм.
Молекулярные цепи оказываются правильно упакованными в среднем на участке длиной 15-17 нм, а затем следует участок "разрыхления" длиной 2,5-3,0 нм. Внутри аморфных областей имеются пустоты, поры размером 0,5-1,0 нм. Архитектоника целлюлозного волокна следующая: 10-12 пачек агрегируются в первичную элементарную фибриллу, 10-12 элементарных фибрилл - во вторичную фибриллу, 10-12 вторичных фибрилл - в микрофибриллу, 10-15 микрофибрилл - в фибриллы. Среднестатистические размеры элементарной фибриллы 20 × 20 нм.
155

Рис. 3.19. Схематическое изображение разреза кристаллической области пачки макромолекул целлюлозы (1 - поперечное сечение полимерной цепи)
Между пачками остаются поры, пустоты размером до 1-2 нм. Вторичные фибриллы имеют среднее сечение 80-120 нм. Размеры пор 5-6 нм. Сечение микрофибрилл - около 0,3 мкм, а размеры пор - до 10 нм.
Укрупнение структурных элементов идет по длине. Длина элементарной фибриллы - около 30 нм, а макрофибриллы - 2-3 мкм. Видимые в электронном микроскопе надмолекулярные структуры целлюлозы представляют собой частицы со степенью асимметрии 1:10 - 1:15.
Целлюлозный материал - это не монолит, а сложный ансамбль молекул, плотность упаковки которых на различных этапах агрегации различна. Наличие пор в материале определяет реакционную способность целлюлозного волокна, его накрашиваемость и т.п.
Структурообразование в белковом волокне (фиброин, кератин) протекает принципиально так же, как и в целлюлозном волокне. Равновесной конформацией макромолекул кератина является α-спираль. Присутствие цистина обусловливает возникновение между полимерными цепями кератина химических связей - дисульфидных (цистиновых) связей. Три макромолекулы кератина ассоциируются в пачку (протофибриллу), 11 протофибрилл - в микрофибриллу, и т.д. Наличие областей с различной плотностью упаковки на разных стадиях надмолекулярной организации определяет структурные различия шерстяных волокон.
156
