6.3.8. Химические реакции полипептидов
Полипептиды, являющиеся стереорегулярными сополимерами, способны, аналогично другим гетероцепным полимерам, как к реакциям деполимеризации (гидролитической деструкции), так и к разнообразным полимераналогичным превращениям. Специфичность строения макромолекул белков обусловливает возможность протекания сопряженных процессов деструкции цепей и полимераналогачных превращений. Кинетика химических реакций в белках определяется не только реакционной способностью тех или иных функциональных групп, но и всеми структурными уровнями полимерного субстрата.
Гидролитическая деструкция
Полипептиды под действием кислотных и щелочных агентов способны к гидролитической деструкции. Гидролиз белков кипящими растворами разбавленных кислот (НСl, H2SO4) приводит к практически полному (до 96-98%) распаду полимерного субстрата на элементарные аминокислоты:
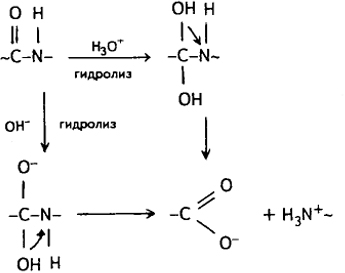
Процесс гидролитической деструкции протекает стохастически в такой последовательности:
белок → альбумин → пептон → полипептид → олигопептиды → дипептиды → аминокислота.
357
Заключительная стадия процесса "дипептид → аминокислота" в некоторых случаях реализуется с трудом, что обусловлено возможностью изменения химической природы аминокислотных звеньев, например:

Наиболее подверженными гидролитическому распаду оказываются пептидные связи у Ser и Thr.
Вопрос. Скорость гидролиза кератина возрастает, если подвергнуть шерсть обработке пероксидом водорода. Какова причина этого явления?
Ответ. Состав белка кератина характеризуется сравнительно высоким содержанием цистина, Cys-Scy (до 8-12%). При воздействии на меж- и внутримолекулярные дисульфидные связи H2O2 или другого окислителя происходит их разрыв с образованием сульфоцистеиновых групп:

Эти группы способны интенсивно гидратироваться, что приводит к значительному набуханию полимерного субстрата в реакционной среде, облегчая доступ гидролизующего агента к связям ~CONH-, ускоряя тем самым гидролитический распад белка.
Оценка стабильности пептидных связей по отношению к гидролитическому воздействию может быть проведена известным в органической химии "правилом шести", сущность которого заключается в том, что стерический (экранирующий) эффект заместителей проявляется тем отчетливей, чем больше атомов С находится в положении 6 относительно двойной связи:

Действительно, полипептидная цепь -Ala-Leu-Lys- под влиянием гидролизующих агентов будет преимущественно разрушаться по связи Ala-Leu.
Следует отметить, что при достаточно жестких гидролитических обработках некоторые боковые радикалы претерпевают глубокие химические изменения. Так, при длительном нагревании белка в 6 н. растворе НСl при 110°С разрушаются Try, Cys, Thr. Еще бо́льшие изменения полимерного субстрата происходят в результате щелочного гидролиза. Так, при нагревании белка
358
в 0,1 н. растворе NaOH при 100°С разрушаются Arg, Thr, Cys.
Вопрос. Для укрепления человеческих волос часто используют реакции превращения цистиновой (дисульфидной) связи в лантиониновую. Каков механизм превращения? Как изменяется строение макромолекулы белка?
Ответ. Под влиянием щелочных обработок при повышенной температуре происходит распад дисульфидных связей в полимерном субстрате кератина - основного белка человеческого волоса, при этом деструкции полимерной цепи практически не наблюдается:

Таким образом, при щелочном гидролизе может происходить не только деструкция полимерной цепи, но и изменение первичной структуры белка, о чем свидетельствуют данные, приведенные на рис. 6.11.
При кислотном гидролизе в оптимальных условиях происходит исчерпывающий распад полипептидной цепи до элементарных аминокислот.
Процессам гидролиза белков предшествует гидратация, вызывающая их набухание и растворение. Гидратация ионизированных групп полимерного субстрата обусловлена ориентацией диполей молекул воды в электрическом поле ионизированной группы (-СОO-; -NH
и др.), а гидратация полярных заместителей - ориентацией молекул воды в результате образования водородных связей. Молекулы гидратно-связанной белком воды можно представить в виде монослоя вокруг ионизированных и полярных групп полипептида, в то время как гидрофобные радикалы
359
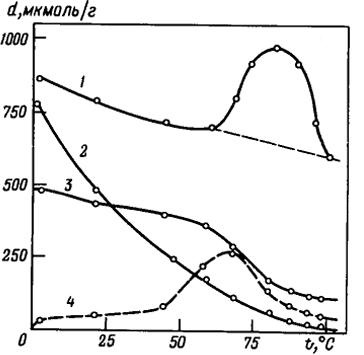
Рис. 6.11. Изменение аминокислотного состава кератина после омыления 0,1 н. раствором NaOH в течение 1 ч при различных температурах (d - содержание аминокислотных звеньев). Содержание связей:
1 - Ser; 2 - Cys; 3 - Thr; 4 - LAN
остаются свободными от воды, реализуя эффекты "гидрофобных взаимодействий". Таким образом, топография гидратного слоя на макромолекуле белка выражается рядом своеобразных "островков". Обычно количество связываемой белком воды составляет 15-35% (масс.). Ориентация диполей воды в результате взаимодействия с полипептидами приводит к уменьшению энтропии системы ΔSг. Термодинамические характеристики процесса гидратации некоторых фибриллярных белков приведены в табл. 6.9.
При дальнейшем увеличении содержания воды наблюдается более интенсивное возрастание значений ΔGг, нежели ΔHг, что свидетельствует об увеличении ΔGг системы.
Таким образом, различная доступность связей ~CONH- гидролитическому распаду определяется преимущественно особенностями первичной структуры макромолекулы. Это явление позволяет решать задачи выбора специфических деструктирующих реагентов, способных селективно разрывать пептидные связи между определенными аминокислотными звеньями. Наиболее подходящими в этом отношении являются гидролитические ферменты. Например, фермент трипсин разрывает связь ~CONH- практически исключительно между Arg и Lys. Другой фермент, химотрипсин, разрывает пептидные связи преимущественно между звеньями, имеющими ароматические ядра (например, между Тyr и Phe).
Таблица 6.9. Термодинамические свойства системы фибриллярный белок - вода, отнесенные к 298 К (в кДж/основомоль)

360
