6.3.3. Вторичная структура белков
Вторичная структура белковой молекулы - это конформация участков полипептидной цепи. Линейный полимер, первичная структура которого включает много "шарнирных" групп и взаимодействие между боковыми радикалами в котором не очень велико, образует статистический клубок. Он не обладает определенной трехмерной структурой или формой, так как она постоянно изменяется под действием микроброуновского движения. Однако вследствие взаимодействия боковых заместителей аминокислотных звеньев макромолекулы белка способны свертываться в более плотный, чем статистический, клубок, в результате чего возникает компактная глобулярная структура белковой макромолекулы.
Полипептидные цепи способны образовывать α-спиральную конформацию (рис. 6.10). Такая конформация характеризуется максимальным насыщением водородных связей вдоль оси спирали. Боковые заместители аминокислотных звеньев направлены наружу и находятся вне спирали. Дополнительным фактором, фиксирующим α-спиральную конформацию макромолекулы белка, является образование внутрицепных дисульфидных (цистиновых), сложноэфирных и солевых связей. Возникновение двойных и тройных спиралей обусловлено интенсивными межмолекулярными взаимодействиями между ними. Такие спиральные одно- и многоцепочечные макромолекулы являются примером стержнеобразных жестких цепей, характеризующихся fФ < 0,63.
β-Структура отвечает растянутым участкам полипептидной цепи и характерна для структуры плоских складчатых слоев белкового
344
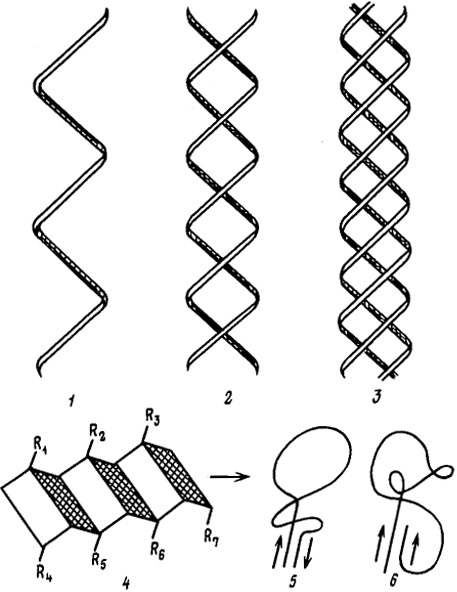
Рис. 6.10. Некоторые конформации макромолекул белков:
1 - α-спираль; 2 - двойная спираль; 3 - тройная спираль (фиброин, коллаген); 4 - β-складчатая структура; 5 - антипараллельное направление β-структуры; 6 - параллельное направление β-структуры
субстрата (см. рис. 6.10). Такая конформация полипептидных цепей предопределяет возможность параллельной и антипараллельной укладки макромолекул и возникновение между ними достаточно прочных межмолекулярных контактов. Остов цепи не лежит в одной плоскости: вследствие изгибов при Сα-атомах слой белкового субстрата оказывается слегка волнистым. Боковые заместители аминокислотных звеньев располагаются перпендикулярно плоскости слоев. В фибриллярных белках полипептидные цепи в β-структурах упакованы антипараллельно. Таким образом, макромолекулы белка способны к обратимым конформационным переходам:

345
В результате таких переходов наблюдается изменение характеристической вязкости, оптической плотности, коэффициента седиментации и других гидродинамических и оптических свойств растворов белков.
Переходы спираль - клубок происходят в растворах белков при нагревании, при изменении рН и концентрации солей.
Фиксация конформаций макромолекул (вторичной структуры) белка происходит в результате различных внутри- и межцепных взаимодействий. Ниже приведена схема внутри- и межцепных взаимодействий в макромолекуле белка [связи: 1 - водородные и диполь-дипольные, 2 - "гидрофобные", 3 - ковалентная дисульфидная, 4 - ковалентная сложноэфирная, 5 - ионная ("солевая")]:
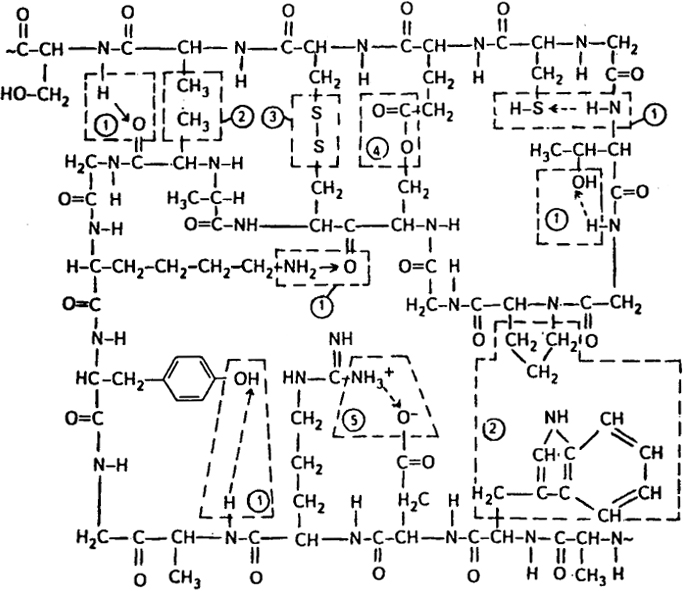
В результате межцепных и внутримолекулярных контактов в белках могут реализоваться:
ковалентные связи типа
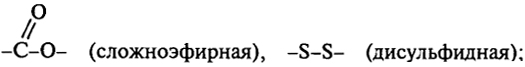
346
солевая

водородные
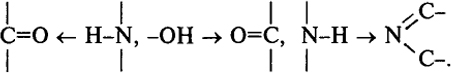
Полипептидная цепь включает участки, имеющие различную конформацию. Поэтому в процессах структурообразования реализуется преимущественно лишь одна из возможных конформаций.
347
