6.3.2. Первичная структура белков
Структурной основой белков является полипептидная цепь. Геометрические параметры пептидной связи приведены на рис. 6.8, а. Все атомы пептидной связи находятся преимущественно в одной плоскости. Уровни структурной организации белков описываются аналогично другим полимерам. При жесткой пептидной связи и фиксированных геометрических параметрах конформация полипептидной цепи описывается двухгранными углами Фi и φi при Сα-атомах (рис. 6.9). Вращение вокруг амидной связи C-N фактически заторможено. Пептидная связь способна к таутомерным переходам по схеме
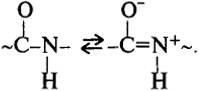
Планарность этой связи и возможность возникновения диполя обусловливают ее жесткость. Жесткая пептидная связь существенно ограничивает возможность конформационных переходов в макромолекуле. Боковые радикалы аминокислотных звеньев создают дополнительные стерические затруднения для конформационных переходов. Звенья Gly лишены бокового радикала и не проявляют асимметрии при Сα-атоме. В связи с этим данные звенья в полипептидной цепи играют роль своеобразного шарнира, позволяя остаткам Gly обеспечивать возможность конформационных переходов. Однако содержание Gly в полимерных цепях ограничено: его избыток привел бы к резкому возрастанию гибкости макромолекул. Важной особенностью строения полипептидной цепи является тот факт, что все полярные и неполярные боковые радикалы отделены от Сα-атома группой СН2, что обеспечивает увеличение конформационных возможностей полимерной цепи. Остов полимерной цепи образует цепочка атомов
-N-C-C-N-C-C-N-C-C-N-C-C-
Боковые заместители аминокислотных звеньев направлены либо внутрь, либо к поверхности белковой молекулы. Неполярные боковые радикалы Val, Ile и Leu разветвлены (см. табл 6.7), что ограничивает их внутреннюю подвижность. Подвижность ароматических циклов в Phe незначительна. Неполярный Pro является специфическим остатком, образующим циклическое звено в полимерной цепи, в результате чего конформационные возможности макромолекулы белка ограничиваются. К тому же Pro фиксирует двухгранный угол Фi между N и Сα в узком интервале ±20 град. Try характерен самым объемным боковым радикалом. Его небольшая полярность обусловлена индольным гетероциклом. Следует отметить, что все самые крупные боковые радикалы Val, Ile, Leu, Phe, Pro, Try, а также Met располагаются преимущественно внутри глобулизированной белковой молекулы.
341
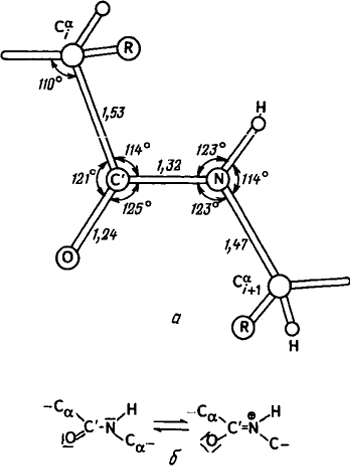
Рис. 6.8. Параметры пептидной связи:
а - валентные углы и межатомные расстояния в транспептидной связи; б - резонансные формы пептидной связи.
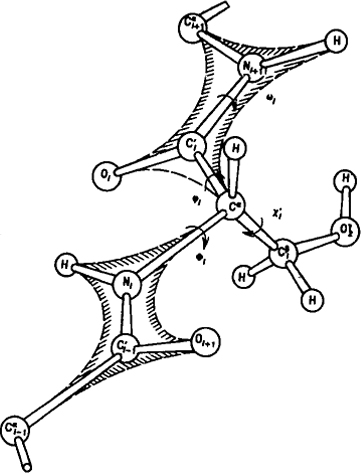
Рис. 6.9. Возможности конформационных переходов в полипептидной цепи. На схеме приведен аминокислотный остаток серина (Ser). Все двухгранные углы в боковом радикале обозначены буквой χi
Типичными полярными и нейтральными боковыми радикалами обладают Ser, Cys, Thr, Arg, Gln и Thr. Они способны образовывать внутри- и межцепные водородные связи. Эти звенья могут располагаться в макромолекуле белка как внутри, так и на поверхности глобулы. Звенья Asp и Glu, как правило, находятся также на поверхности частиц белков. Формирование вторичной структуры белка зависит как от особенностей первичной структуры, так и от внешних (влажность, рН, температура) условий.
Вопрос. Натуральный шелк производится в основном путем физико-химических обработок коконов тутового шелкопряда Bombyx mori. Коконы, формуемые гусеницами этого насекомого, представляют собой природную композиционную оболочку, построенную из прочных фиброиновых волокон ("стержней"), связанных между собой другим линейным белком - серицином (табл. 6.8). Для защиты от болезнетворных бактерий, обеспечения газо- и водообмена с окружающей средой поверхность нитей, из которых построен кокон, покрыта тонкой полупроницаемой белково-липидной оболочкой. Коконная нить на 97-98% состоит из высокоориентированных фибриллярных белков:
342
Таблица 6.8. Композиционный состав некоторых белков
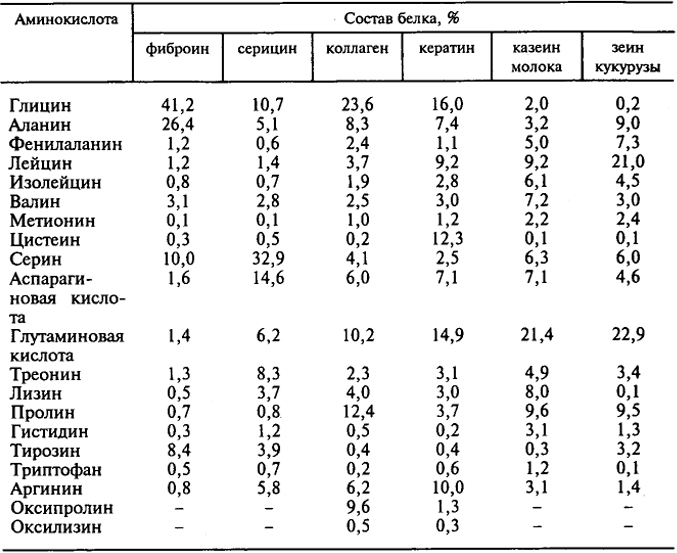
фиброина и серицина. Однако количественное разделение белковых компонентов этой нити практически невозможно, так как отсутствует четкая граница в первичной структуре этих белков.
При производстве натурального шелка коконная нить (кокон) подвергается обработке горячей водой в присутствии поверхностно-активных веществ (ПАВ), в результате чего получают полупродукт - шелк-сырец. Последующая дополнительная гидротермическая обработка ("отварка") его приводит к получению текстильного натурального шелка, содержащего 4-6% остаточного серицина. Исходя из особенностей первичной структуры фиброина и серицина (см. табл. 6.8), расскажите, какие основные физико-химические процессы происходят при получении натурального шелка.
Ответ. В процессе гидротермических обработок ("размотка" коконов, "отварка" шелка-сырца) происходит количественное удаление из нити жиро-восковых веществ и значительной части водорастворимого белкового компонента - серицина, первичная структура которого характеризуется увеличенным содержанием аминокислотных звеньев с гидрофильными боковыми радикалами: Ser, Asp, Glu, Thr, Lis. Экстракция серицина происходит в условиях интенсивного набухания полимерного субстрата.
В результате таких обработок фиброиновый компонент нити - натуральный шелк - оказывается существенно более гидрофобным, нежели исходная коконная нить, и после сушки приобретает необходимые текстильно-технологические характеристики.
343
При полном удалении воды из нити натурального шелка (например, в результате криогенной сушки) он становится хрупким материалом.
Огромное число взаимных сочетаний α-аминокислотных звеньев в полипептидной цепи, обусловливающих первичную структуру белка, предопределяет возможность существования очень большого разнообразия белков- и специфичность их функций. Однако первичная структура белка, обладающая специфическими функциональными свойствами (например, фибриллярные белки), в процессе биосинтеза воспроизводится достаточно точно, что обусловливает возможность жизнедеятельности организмов. Ранее уже отмечалось, что конформационные переходы в полипептидной цепи могут осуществляться в основном в результате вращения вокруг СН2-группы Gly, играющей роль шарнира.
344
